Что являются переносчиками газов кислорода и углекислого газа. Транспорт газов кровью. Транспорт кровью углекислого газа и его связь с кислотно-щелочным состоянием крови
Транспорт газов кровью. Транспорт О2 и СО2. Кривая диссоциации Нв.
Транспортная функция крови заключается в переносе всех необходимых для жизнедеятельности организма веществ (питательных веществ, газов, гормонов, ферментов, метаболитов).
Дыхательная функция состоит в доставке кислорода от легких к тканям и углекислого газа от тканей к легким.
Транспорт газов кровью – в организме кислород и углекислый газ транспортируются кровью. Кислород, поступающий из альвеолярного воздуха в кровь, связывается с гемоглобином эритроцитов, образуя так называемый оксигемоглобин, и в таком виде доставляется к тканям.
Кислород через тонкие стенки альвеол и капилляров поступает из воздуха в кровь, а углекислый газ из крови в воздух. Диффузия газов происходит в результате разности их концентраций в крови и в воздухе. Кислород проникает в эритроциты и соединяется с гемоглобином, кровь становится артериальной и направляется в ткани. В тканях происходит обратный процесс: кислород за счет диффузии переходит из крови в ткани, а углекислый газ, наоборот, переходит из тканей в кровь.
Кривая диссоциации оксигемоглобина — это зависимость насыщения гемоглобина кислородом (измеряется процентным отношением оксигемоглобина к общему количеству гемоглобина).
Газообмен между кровью и тканями. Нарушение газообмена в тканях.
Газообмен в тканях –четвертый этап дыхания, в результате которого кислород из крови капилляров поступает в клетки, а углекислый газ из клеток в кровь. Фактором, способствующим газообмену в тканях, является, также как и в легких, разность парциальных давлений газов между кровью и межтканевой жидкостью, омывающей все клетки и ткани.
Насколько клетки интенсивно поглощают кислород, настолько же интенсивно они вырабатывают углекислый газ. Напряжение углекислоты в клетках достигает 50 — 60 мм. рт. ст. Эта углекислота непрерывно переходит в межтканевую жидкость, а оттуда в кровь, делая кровь венозной.
Следствием таких нарушений газообмена может явиться гипоксия, кислородное голодание тканей. Гипоксия – это недостаток кислорода.
Газообмен в легких, состав вдыхаемого альвеолярного, выдыхаемого воздуха.
Как атмосферный воздух, так и альвеолярный необходимо воздух представляют собой смесь газов, содержащую О2 , СО2 , N, и инертные газы. Определенное количество дыхательных газов содержится и в крови, поскольку она является их переносчиком. Парциальное давление того или иного газа в крови, ровно как и в любой другой жидкости, принято называть парциальным напряжением. Газообмен между альвеолярным воздухом и кровью капилляров (второй этап дыхания) осуществляется путем диффузии, благодаря разности давлений О2 и СО2. Тот воздух, который мы вдыхаем, т.е. атмосферный воздух, имеет более или менее постоянный состав: он содержит
0,03% углекислого газа
79,03% азота.
Выдыхаемый воздухобеднен кислородом и насыщен углекислотой. В среднем выдыхаемый воздух содержит
По сравнению с атмосферным воздухом, альвеолярный воздух содержит
Состав альвеолярного воздуха относительно стабилен, так как при спокойном дыхании в альвеолы поступает всего 350 мл свежего воздуха, что составляет лишь 1/7 того воздуха, который содержится в легких после обычного выдоха. Данный воздух находится в альвеолах и обеспечивает потребление кислорода для обменных процессов в капиллярах легких.
Такая же небольшая порция альвеолярного воздуха удаляется при выдохе, что способствует стабилизации его состава.
Регуляция дыхания. Нервно-регуляторная и гуморальная регуляция дыхания.
Внешнее дыхание-это обмен воздуха между альвеолами легких и внешней средой, которое осуществляется в результате ритмического дыхания движения грудной клетки ,вызывающих чередование актов вдоха и выдоха.
Главная цель внешнего дыхания – поддержание оптимальный состав артериальной крови. Основной способ для достижения этой цели – регулирование объема легочной вентиляции путем изменения частоты и глубины дыхания. Какие же механизмы обеспечивают приспособление дыхания к меняющимся потребностям организма? Организм располагает двумя регуляторными системами – нервной и гуморальной. Последняя представленациркулирующими в крови гормонами и метаболитами, которые могут влиять на дыхание.
Регуляция дыхания –называется процесс управления вентиляцией легких,направленный на поддержание дыхательных констант и приспособления дыхания к условиям изменяющейся внешней среды.
Следовательно, для осуществления дыхательных движений нужен продолговатый мозг и тот отдел спинного мозга, который посылает двигательные нервы к дыхательным мышцам.
Дата добавления: 2018-08-06 ; просмотров: 520 ;
Транспорт кислорода и углекислого газа кровью
ОСНОВНЫЕ ПОЛОЖЕНИЯ ФИЗИОЛОГИИ И ПАТОФИЗИОЛОГИИ
ВНЕШНЕГО ДЫХАНИЯ
Основной функцией системы внешнего дыхания является оксигенация крови и удаление углекислого газа. Внешнее дыхание можно разделить на два этапа: вентиляция легких и газообмен в них. Вентиляция — это процесс вдоха и выдоха. Процесс вдоха обеспечивается сокращением дыхательных мышц, основной мышцей вдоха является диафрагма. Сокращение дыхательных мышц приводит к уменьшению внутриплеврального давления на 8—10 см вод. ст. ниже атмосферного давления за счет увеличения объема грудной клетки. Вследствие этого увеличивается объем легких, а давление в альвеолах понижается на 1—2 см вод. ст. ниже атмосферного давления, и воздух на вдохе поступает у альвеолы. Разницу между внутриплевральным и внутриальвеолярным давлением называют транспульмональным давлением, за счет которого и происходит расширение легких.
Для непосредственного сокращения дыхательных мышц необходима импульсация из дыхательного центра, нейроны которого расположены в ретикулярной формации продолговатого мозга. Нервные импульсы, которые генерируют нейроны дыхательного центра, проходят по проводящим путям спинного мозга, где расположены мотонейроны дыхательных мышц, далее направляются по нервным волокнам к нервно-мышечным синапсам и затем стимулируют сокращение дыхательных мышц. Мотонейроны диафрагмы расположены в CI—CV сегментах спинного мозга (респираторный тракт), где образуют диафрагмальные нервы, которые являются двигательными нервами диафрагмы. Мотонейроны дыхательных межреберных мышц расположены посегментарно в спинном мозге; импульсы от них, в основном, идут по межреберным нервам.
Вентиляция легких направлена на поддержание нормального состава альвеолярного воздуха. Каков нормальный состав альвеолярного воздуха?
Для освещения этого вопроса необходимо остановиться на определении парциального давления газа в смеси газов. Согласно закону Дальтона, смесь газов образует на стенке закрытой емкости давление, которое равняется сумме парциальных давлений всех газов смеси, а парциальное давление каждого газа в смеси прямо пропорционально его концентрации в смеси. Таким образом, если в атмосферном воздухе концентрация кислорода составляет 20,91 %, а атмосферное давление на уровне моря — 760 мм рт. ст., то парциальное давление кислорода в атмосфере будет составлять около 1/5 от атмосферного давления, или 150 мм рт. ст. (20 кПа).
Давление альвеолярного воздуха равно атмосферному при температуре тела 37 0 С. В нем на водяные пары припадает 47 мм рт. ст., на все другие газы остается 713 мм рт. ст. Вследствие того, что азот является биологически инертным газом, его концентрация в альвеолах такая же, как и в атмосфере, а именно — 79 %. Таким образом, на кислород и углекислый газ остается около 21 % от 713 мм рт. ст. При условиях нормальной вентиляции парциальное давление углекислого газа в альвеолярном воздухе (РAСО2) составляет 40 мм рт. ст. (5,3 кПа), тогда:
где АлД — альвеолярное давление, которое равно атмосферному и составляет с поправкой на дыхательный коэффициент немногим более 100 мм рт. ст., или 13,3 кПа.
Основным показателем адекватности вентиляции легких принято считать РAСО2.
Следующим этапом внешнего дыхания является газообмен в легких. Обмен кислорода и углекислого газа между альвеолярным воздухом и кровью легочных капилляров осуществляется путем диффузии через альвеолокапиллярную мембрану. Согласно закону диффузии Фика, скорость диффузии (M/t) прямо пропорциональна разности парциальных давлений газов с обеих сторон мембраны (ΔР), площади диффузии (S, в норме — альвеолярная поверхность), коэффициенту диффузии (k), коэффициенту растворимости газа в жидкости (ά, поскольку в легочном интерстиции и на поверхности альвеол присутствует жидкость) и обратно пропорциональна толщине мембраны (х):
M/t = (ΔP x S x k x ά)/x.
ΔР для кислорода равна 60—70 мм рт. ст., углекислого газа — 6 мм рт. ст. Несмотря на это, а также на значительную величину коэффициента диффузии для кислорода, за счет того, что коэффициент растворимости для углекислого газа намного больше, он диффундирует через альвеолокапиллярную мембрану более чем в 20 раз быстрее кислорода. Благодаря широкой поверхности диффузии (альвеолярная поверхность в среднем составляет 80 м 2 ) резервы диффузии в легких немалые, поэтому в клинической практике нарушения диффузии, как основной фактор нарушения газообмена, имеют принципиальное значение практически только при отеке легких.
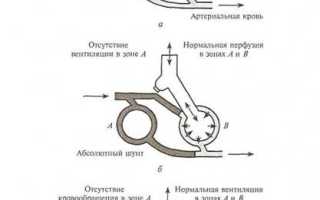
Кроме диффузии, для нормального газообмена в легких необходимо нормальное соотношение между альвеолярной вентиляцией и легочной перфузией (VA/QC), которое в норме составляет 0,8—1,0. При условии увеличения VA/QC вентилируются альвеолы, которые не перфузируются, поэтому развивается гипервентиляция с уменьшением РAСО2 (гипокапния). При условии уменьшения VA/QCразвивается гипоксемия (уменьшение РО2 в артериальной крови). Особо опасно уменьшение VA/QC до 0, когда перфузия альвеол сохранена, а вентиляция отсутствует (шунтирование крови справа налево – Qs/Qt, где Qs- венозная примесь, Qt – сердечный выброс), и венозная кровь без оксигенации и отдачи углекислого газа поступает в легочные вены. Виды нарушений вентиляционно-перфузионных соотношений приведены на рис. 1.
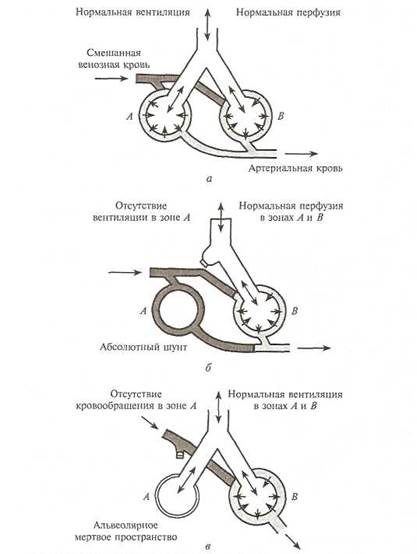
Рис. 1. Виды нарушений вентиляционно-перфузионных соотношений. Три модели соотношения вентиляции к перфузии в легких: а — норма, б — шунт, в — альвеолярное мертвое пространство.
Если Qs/Qt превышает 10 % сердечного выброса, то возникает гипоксемия, если 40 % — гиперкапния. Наиболее часто увеличение внутрилегочного Qs/Qt в клинической практике возникает при ателектазах легких, пневмонии, ОРДС.
Транспорт кислорода и углекислого газа кровью
1. Вентиляция легкихобеспечивает доставку кислорода из воздуха (РiО2 = 158 мм рт. ст.) к альвеолярному газу (РAО2 = 105-110 мм рт. ст.), выведение СО2 из альвеолярного газа (РAСО2 = 40 мм рт. ст.) в атмосферу.
Вентиляция у здорового человека приспособлена к метаболическим потребностям таким образом, что напряжение углекислоты в альвеолярном воздухе и артериальной крови (РаСО2) поддерживается на уровне 37-40 мм рт.ст., а напряжение кислорода в артериальной крови (РаО2) – в пределах 95-98 мм рт.ст.
Вентиляция легких зависит от дыхательного объема (в физиологических условиях 400-500 мл) и частоты дыхания (в норме 12-16 в ми). Произведение дыхательного объема на частоту дыхания (ЧД) составляет минутный объем дыхания (МОД).
В процессе дыхания не весь вдыхаемый воздух участвует в газообмене. Часть его, около 1 /3 МОД, остается в мертвом пространстве(ОМП), которое включает в себя верхние дыхательные пути (глотка, трахея, бронхи) и невентилируемые альвеолы. Только 2 /3 МОД достигает альвеол, что составляет минутную альвеолярную вентиляцию (МАВ). Зависимость между МОД и МАВ выражается формулой: МАВ = МОД – ОМП × ЧД. Следует отметить, что МАВ более важный показатель внешнего дыхания, чем МОД. Так при одышке свыше 30 в мин, несмотря на большой МОД альвеолярная вентиляция обычно снижается. При более низком МОД и замедлении дыхания МАВ может увеличиваться. Например, при МОД – 8000 мл, ЧД – 40 в мин и ОМП – 150 мл МАВ = 8000 – (150 × 40) = 2000 мл, а при МОД – 6000 мл, ЧД – 10 в мин и ОМП – 150 мл МАВ = 6000 – (150 × 10) = 4500 мл.
2.Газообмен в легкихобеспечивает поступление кислорода из альвеолярного газа в артериальную (капиллярную) кровь (РАО2 = 100 мм рт. ст.), СО2 удаляется из венозной крови легочных капилляров (PvCO2 = 46 мм рт. ст.) в альвеолярный газ.
3.Малый круг кровообращенияобеспечивает доставку кислорода из легких по легочным венам к левому предсердию, СО2 транспортируется из правого желудочка к альвеолам.
Большой круг кровообращенияобеспечивает доставку кислорода по артериям к капиллярам (РСО2 уменьшается с 100 мм рт. ст. до 40), СО2 — из капилляров (РсСО2 — с 40 до 46 мм рт. ст.) в легкие.
Кислород переносится к тканям в виде его соединения с гемоглобином эритроцита и в незначительном количестве растворенным в плазме. Так как 1 г гемоглобина способен связать 1,34 мл О2, кислородная емкость крови при нормальном содержании гемоглобина (150 г/л) составляет приблизительно 20 мл О2 на 100 мл крови, то есть 20 об%. Кроме того, 100 мл крови переносят 0,3 мл кислорода, растворенного в плазме. Даже минимальное количество кислорода, переносимого плазмой, может играть важную роль при возрастании его парциального давления. Повышение РаО2 на 1 мм рт.ст. (0,13 кПа) увеличивает содержание кислорода в плазме на 0,003 об%. Таким образом, обычно, в 100 мл крови кислорода содержится около 2 об% (760 × 0,003), а в барокамере при давлении 3 атмосферы около 6 об%. Этого достаточно для обеспечения организма кислородом при выраженной анемии.
У здорового человека не весь гемоглобин связывается с кислородом. Это обусловлено физиологическим артерио-венозным шунтированием в легких, при котором часть крови проходит через невентилируемые альвеолы. Поэтому насыщение (сатурация) крови кислородом (SаО2) в норме соответствует 96-98%, а не 100%. Величина SаО2 зависит и от парциального напряжения кислорода в крови (РаО2), которое в норме равно 96-98 мм рт.ст. (42,8-43,1 кПа). Между изменениями РаО2 и SаО2 нет полного соответствия, так как SаО2 даже при дыхании 100% кислородом под давленим 2-3 атмосферы может достигнуть только 100%, а РаО2 при этом повысится до 400-600 мм рт.ст. (53-80 кПа), то есть в 3-4 раза.
4.Транскапиллярный обмен газов:кислород переходит из капиллярной крови в интерстициальную жидкость, а затем — в клетки, где в митохондриях, благодаря механизмам тканевого дыхания (НАД, ФАД, цитохромы, цитохромоксидаза), окисляет водород с образованием воды и энергии, которая аккумулируется в АТФ; СО2, образующийся в цикле Кребса, переходит в капиллярную кровь.
Обмен кислорода на тканевом уровне обеспечивается сохранением градиента давления, что приводит к переходу О2 из тканевых капилляров путем диффузии к месту утилизации (митохондрии клеток).
При недостатке кислорода организм компенсирует его дефицит, переключаясь на менее эффективный тип дыхания – анаэробный.
В упрощенной схеме оба пути можно представить следующим образом. Анаэробный путь: глюкоза – пировиноградная кислота – молочная кислота + 2 молекулы АТФ (16 кал. свободной энергии). Аэробный путь: глюкоза – пировиноградная кислота – СО2 + Н2О + 38 молекул АТФ (304 кал. свободной энергии).
Следовательно, большая часть проблем реанимации связана с необходимостью поддержания напряжения О2 в клетках на уровне, способствующем синтезу АТФ путем аэробного метаболизма. Клеточную гипоксию можно определить как состояние, при котором аэробный метаболизм нарушен.
Углекислота транспортируется кровью в трех основных видах – в растворенном, с бикарбонатом и в соединении с белками (главным образом с гемоглобином) в форме карбаминовых соединений. Если альвеолярная вентиляция становится недостаточной для элиминации выработанной организмом углекислоты, РаСО2 повышается (возникает гиперкапния).
Таким образом, благодаря системе внешнего дыхания кислород поступает в кровь, а СО2 выводится из нее; далее сердце перекачивает кровь, насыщенную кислородом, к тканям, а кровь, насыщенную углекислым газом, — к легким.
Транспорт кислорода (DO2) зависит от сердечного индекса (СИ) и содержания кислорода в артериальной крови (CaO2).
где: k — коэффициент растворимости кислорода (0,031 мл/мм рт. ст./л), G — константа Гюфнера (равна количеству кислорода в мл, которое может присоединить 1 г гемоглобина; в среднем составляет 1,36 (1,34—1,39) мл/г).
При условии СИ = 2,5—3,5 л/мин/м 2 , транспорт кислорода составляет: DO2 = 520-720 мл/мин/м 2 .
Необходимо отметить, что множество патологических состояний, которые нуждаются в оказании неотложной медпомощи, сопровождаются дефицитом доставки и потребления кислорода, что обусловлено недостаточностью дыхания, нарушением кровообращения или анемией. В зависимости от механизма нарушений транспорта кислорода к тканям различают несколько видов гипоксии.
Кроме своей основной, дыхательной, функции легкие осуществляют нереспираторные (недыхательные) функции механического и метаболического характера, которые связывают легкие с другими системами организма.
Нереспираторные (недыхательные) функции легких:
· защитная – легкие задерживают до 90% вредных механических и токсических продуктов (частицы диаметром болем 2 мкм), которые поступают из окружающей среды (важную роль при этом играют слизь дыхательных путей, которая содержит лизоцим и иммуноглобулины, макрофаги и альвеолоциты I та II типа);
· очистительная (фильтрационная) – легкие очищают кровь от механических примесей (агрегатов клеток, капель жира, мелких тромбов, бактерий, крупных атипичных клеток), которые задерживаются в них и подвергаються деструкции и метаболизму;
· фибринолитическая и антикоагулянтная – улавливание легкими тромбов, поддержание фибринолитической и антикоагулянтной активности крови;
· деструкция белков и жиров – легкие богаты протеолитическими и липолитическими ферментами; в легких продуцируется сурфактант – комплекс липопротеидов, который способствует стабильности альвеолярной ткани;
· участие в водном балансе – легкие удаляют за сутки (посредством перспирации) около 500 мл воды, поддерживая нормальную осмолярность крови и тканей удалением СО2 и соответственным изменением уровня осмотически активних карбонатов (15-30 мосмоль/сутки); вместе с тем, различные жидкости могут активно всасываться в легких, например адреналин уже через 30 с определяется в крови;
· избирательная деструкция биологически активных веществ (серотонин, гистамин, ангиотензин, ацетилхолин, норадреналин, кинины и простагландины), которые, выполнив свою роль в тканях, подлежат удалению из крови;
· детоксикационная функция – в легких осуществляется метаболизм некоторых лекарственных препаратов – аминазина, индерала, сульфаниламидов и др.;
· участие в теплопродукции и теплоотдачи – суточный теплообмен легких в нормальных условиях составляет 350 ккал, а в условиях критического состояния может быть увеличен в несколько раз;
· гемодинамическая функция – легкие являются резервуаром и одновременно прямым шунтом между правой и левой половинами сердца.
В нормальных условиях для выполнения этих функций необходимо не менее 10% общего поглощенного организмом О2. Во время критических состояний это количество возрастает во много раз.
Что являются переносчиками газов кислорода и углекислого газа. Транспорт газов кровью. Транспорт кровью углекислого газа и его связь с кислотно-щелочным состоянием крови
Кровообращение выполняет одну из важнейших функций переноса кислорода от легких к тканям, а углекислого газа — от тканей к легким. Потребление кислорода клетками тканей может изменяться в значительных пределах, например при переходе от состояния покоя к физической нагрузке и наоборот. В связи с этим кровь должна обладать большими резервами, необходимыми для увеличения ее способности переносить кислород от легких к тканям, а углекислый газ в обратном направлении.
Транспорт кислорода.
При 37 С растворимость 02 в жидкости составляет 0,225 мл • л-1 • кПа-1 (0,03 мл/л/мм рт. ст.). В условиях нормального парциального давления кислорода в альвеолярном воздухе, т. е. 13,3 кПа или 100 мм рт.ст., 1 л плазмы крови может переносить только 3 мл 02, что недостаточно для жизнедеятельности организма в целом. В покое в организме человека за минуту потребляется примерно 250 мл кислорода. Чтобы тканям получить такое количество кислорода в физически растворенном состоянии, сердце должно перекачивать за минуту огромное количество крови. В эволюции живых существ проблема транспорта кислорода была более эффективно решена за счет обратимой химической реакции с гемоглобином эритроцитов. Кислород переносится кровью от легких к тканям организма молекулами гемоглобина, которые содержатся в эритроцитах.
Гемоглобин способен захватывать кислород из альвеолярного воздуха (соединение называется ок-сигемоглобином) и освобождать необходимое количество кислорода в тканях. Особенностью химической реакции кислорода с гемоглобином является то, что количество связанного кислорода ограничено количеством молекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет 4 места связывания с кислородом, которые взаимодействуют таким образом, что зависимость между парциальным давлением кислорода и количеством переносимого кислорода с кровью имеет S-образную форму, которая носит название кривой насыщения или диссоциации оксигемоглобина (рис. 10.18). При парциальном давлении кислорода 10 мм рт. ст. насыщение гемоглобина кислородом составляет примерно 10 %, а при Р02 30 мм рт. ст. — 50—60 %. При дальнейшем увеличении парциального давления кислорода от 40 мм рт. ст. до 60 мм рт. ст. происходит уменьшение крутизны кривой диссоциации оксигемоглобина и процент его насыщения кислородом возрастает в диапазоне от 70—75 до 90 % соответственно. Затем кривая диссоциации оксигемоглобина начинает занимать практически горизонтальное положение, поскольку увеличение парциального давления кислорода с 60 до 80 мм рт. ст. вызывает прирост насыщения гемоглобина кислородом на 6 %. В диапазоне от 80 до 100 мм рт. ст. процент образования оксигемоглобина составляет порядка 2. В результате кривая диссоциации оксигемоглобина переходит в горизонтальную линию и процент насыщения гемоглобина кислородом достигает предела, т. е. 100. Насыщение гемоглобина кислородом под влиянием Р02 характеризует своеобразный молекулярный «аппетит» этого соединения к кислороду.
Значительная крутизна кривой насыщения гемоглобина кислородом в диапазоне парциального давления от 20 до 40 мм рт. ст. способствует тому, что в ткани организма значительное количество кислорода может диффундировать из крови в условиях фадиента его парциального давления между кровью и клетками тканей (не менее 20 мм рт. ст.). Незначительный процент насыщения гемоглобина кислородом в диапазоне его парциального давления от 80 до 100 мм рт. ст. способствует тому, что человек без риска снижения насыщения артериальной крови кислородом может перемещаться в диапазоне высот над уровнем моря до 2000 м.
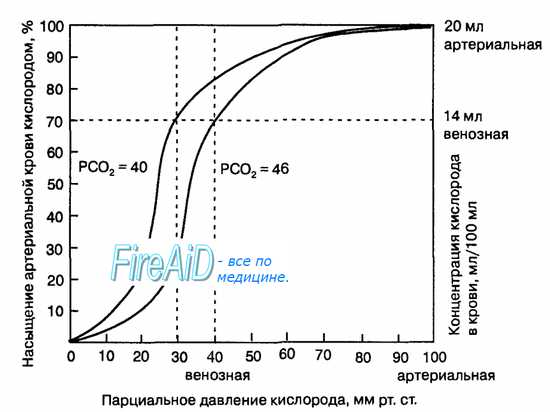 Рис. 10.18. Кривая диссоциации оксигемоглобина. Пределы колебания кривой при РС02 = 40 мм рт. ст. (артериальная кровь) и РС02 = 46 мм рт. ст. (венозная кровь) показывают изменение сродства гемоглобина к кислороду (эффект Ходена).
Рис. 10.18. Кривая диссоциации оксигемоглобина. Пределы колебания кривой при РС02 = 40 мм рт. ст. (артериальная кровь) и РС02 = 46 мм рт. ст. (венозная кровь) показывают изменение сродства гемоглобина к кислороду (эффект Ходена).
Общие запасы кислорода в организме обусловлены его количеством, находящимся в связанном состоянии с ионами Fe2+ в составе органических молекул гемоглобина эритроцитов и миоглобина мышечных клеток.
Один грамм гемоглобина связывает 1,34 мл 02. Поэтому в норме при концентрации гемоглобина 150 г/л каждые 100 мл крови могут переносить 20,0 мл 02.
Количество 02, которое может связаться с гемоглобином эритроцитов крови при насыщении 100 % его количества, называется кислородной емкостью гемоглобина. Другим показателем дыхательной функции крови является содержание 02 в крови (кислородная емкость крови), которое отражает его истинное количество, как связанного с гемоглобином, так и физически растворенного в плазме. Поскольку в норме артериальная кровь насыщена кислородом на 97 %, то в 100 мл артериальной крови содержится примерно 19,4 мл 02.
Транспорт газов кровью
В предыдущих статьях на эту тему вы могли узнать об обмене газов в легких, внешнем дыхании и о строении дыхательных мышц. Сегодня вы узнаете о том, как транспортируются газы в нашей крови.
Переносчиком кислорода от легких к тканям и углекислого газа от тканей к легким является кровь. В свободном (растворенном) состоянии переносится настолько малое количество этих газов, что им смело можно пренебречь при оценке потребностей организма. Для простоты в дальнейшем будем считать, что основное количество кислорода и углекислого газа транспортируется в связанном состоянии.
Транспорт кислорода
Кислород транспортируется в виде оксигемоглобина. Оксигемоглобин — это комплекс гемоглобина и молекулярного кислорода.
Гемоглобин содержится в красных кровяных тельцах — эритроцитах. Эритроциты под микроскопом похожи на слегка приплюснутый бублик, дырку в котором забыли проткнуть до конца. Такая необычная форма позволяет эритроцитам взаимодействовать с окружающей кровью большей площадью, чем шарообразным клеткам (помните — из тел, имеющих равный объем, шар имеет минимальную площадь). А кроме того, эритроцит способен сворачиваться в трубочку, протискиваясь в узкий капилляр и добираясь в самые отдаленные уголки организма.
В 100 мл крови при температуре тела растворяется лишь 0,3 мл кислорода. Кислород, растворяющийся в плазме крови капилляров малого круга кровообращения, диффундирует в эритроциты, сразу же связывается гемоглобином, образуя оксигемоглобин, в котором кислорода 190 мл/л. Скорость связывания кислорода велика — время поглощения диффундировавшего кислорода измеряется тысячными долями секунды. В капиллярах альвеол с соответствующими вентиляцией и кровоснабжением практически весь гемоглобин притекающей крови превращается в оксигемоглобин. А вот сама скорость диффузии газов «туда и обратно» значительно медленнее скорости связывания газов. Отсюда следует второй практический вывод: чтобы газообмен шел успешно, воздух должен «получать паузы», за время которых успевает выровняться концентрация газов в альвеолярном воздухе и притекающей крови, то есть обязательно должна присутствовать пауза между вдохом и выдохом.
Запомните это!
Превращение восстановленного (бескислородного) гемоглобина (дезоксигемоглобина) в окисленный (содержащий кислород) гемоглобин (оксигемоглобин) зависит от содержания растворенного кислорода в жидкой части плазмы крови. Причем механизмы усвоения растворенного кислорода весьма эффективны.
Например, подъем на высоту 2 км над уровнем моря сопровождается снижением атмосферного давления с 760 до 600 мм рт. ст., парциального давления кислорода в альвеолярном воздухе со 105 до 70 мм рт. ст., а содержание оксигемоглобина снижается лишь на 3%. И, несмотря на снижение атмосферного давления, ткани продолжают успешно снабжаться кислородом.
В тканях, требующих для нормальной жизнедеятельности много кислорода (работающие мышцы, печень, почки, железистые ткани), оксигемоглобин «отдает» кислород очень активно, иногда почти полностью. В тканях, в которых интенсивность окислительных процессов мала(например, в жировой ткани), большая часть оксигемоглобина не «отдает» молекулярный кислород — уровень диссоциации оксигемоглобина низкий. Переход тканей из состояния покоя в деятельное состояние (сокращение мышц, секреция желез) автоматически создает условия для увеличения диссоциации оксигемоглобина и увеличения снабжения тканей кислородом.
Способность гемоглобина «удерживать» кислород (сродство гемоглобина к кислороду) снижается при увеличении концентрации углекислого газа и ионов водорода. Подобным же образом действует на диссоциацию оксигемоглобина повышение температуры.
Отсюда становится легко понятным, как взаимосвязаны и сбалансированы относительно друг друга природные процессы. Изменения способности оксигемоглобина удерживать кислород имеет громадное значение для обеспечения снабжения им тканей. В тканях, в которых процессы обмена веществ протекают интенсивно, концентрация углекислого газа и ионов водорода увеличивается, а температура повышается. Это ускоряет и облегчает «отдачу» гемоглобином кислорода и облегчает течение обменных процессов.
В волокнах скелетных мышц содержится близкий к гемоглобину миоглобин. Он обладает очень высоким сродством к кислороду. «Ухватившись» за молекулу кислорода, он уже не отдаст ее в кровь.
Количество кислорода в крови
Максимальное количество кислорода, которое может связать кровь при полном насыщении гемоглобина кислородом, называется кислородной емкостью крови. Кислородная емкость крови зависит от содержания в ней гемоглобина.
В артериальной крови содержание кислорода лишь немного (на 3-4%) ниже кислородной емкости крови. В обычных условиях в 1 л артериальной крови содержится 180-200 мл кислорода. Даже в тех случаях, когда в экспериментальных условиях человек дышит чистым кислородом, его количество в артериальной крови практически соответствует кислородной емкости. По сравнению с дыханием атмосферным воздухом количество переносимого кислорода увеличивается мало (на 3-4%).
Венозная кровь в состоянии покоя содержит около 120 мл/л кислорода. Таким образом, протекая по тканевым капиллярам, кровь отдает не весь кислород.
Часть кислорода, поглощаемая тканями из артериальной крови, называется коэффициентом утилизации кислорода. Для его вычисления делят разность содержания кислорода в артериальной и венозной крови на содержание кислорода в артериальной крови и умножают на 100.
Например:
(200- 120): 200 х 100 = 40%.
В покое коэффициент утилизации кислорода организмом колеблется от 30 до 40%. При интенсивной мышечной работе он повышается до 50-60%.
О транспорте углекислого газа читайте в следующей статье.
Материал подготовил: Atamovich
Источник: Медведев Б.А.
«Животворящее дыхание. Дыхательные практики, которые всегда работают»
Внимание! Этот материал предназначен только для личного использования с целью ознакомления!