

Генетическая рекомбинация при трансформации. Трансдукция у бактерий. Общая и специфическая трансдукция. Использование трансформации и трансдукции для картирования генов. Трансдукция. Неспецифическая трансдукция. Специфическая трансдукция. Абортивная транс
Рекомбинация у бактерий: трансформация, трансдукция, конъюгация
Рекомбинации (обмен генетическим материалом) у бактерий отличаются от рекомбинаций у эукариот:
• у бактерий имеется несколько механизмов рекомбинаций;
• при рекомбинациях у бактерий образуется не зигота, как у эукариот, а мерозигота (несет полностью генетическую информацию реципиента и часть генетической информации донора в виде дополнения);
• у бактериальной клетки-рекомбината изменяется не только качество, но и количество генетической информации.
Трансформация — это обмен генетической информацией у бактерий путем введения в бактериальную клетку-реципиент готового препарата ДНК (специально приготовленного или непосредственно выделенного из клетки-до нора). Чаще всего передача генетической информации происходит при культивировании реципиента на питательной среде, содержащей ДНК донора. Для восприятия донорской ДНК при трансформации клетка-реципиент должна находиться в определенном физиологическом состоянии (компетентности), которое достигается специальными методами обработки бактериальной популяции.
При трансформации передаются единичные (чаще 1) признаки. Трансформация является самым объективным свидетельством связи ДНК или ее фрагментов с тем или иным фенотипическим признаком, поскольку в реципиентную клетку вводится чистый препарат ДНК.
Трансдукция — обмен генетической информацией у бактерий путем передачи ее от донора к реципиенту с помощью умеренных (трансдуцирующих) бактериофагов.
Трансдуцирующие фаги могут переносить 1 или более генов (признаков).
• специфической — переносится всегда один и тот же ген;
• неспецифической — передаются разные гены.
Это связано с локализацией трансдуиируюших фагов в геноме донора:
• в случае специфической трансдукции они располагаются всегда в одном месте хромосомы;
• при неспецифической их локализация непостоянна.
Конъюгация — обмен генетической информацией у бактерий путем передачи ее от донора к реципиенту при их прямом контакте. После образования между донором и реципиентом конъюгационного мостика одна нить ДНК-донора поступает по нему в клетку-реципиент. Чем дольше контакт, тем большая часть донорской ДНК может быть передана реципиенту.
Основываясь на прерывании конъюгации через определенные промежутки времени, можно определить порядок расположения генов на хромосоме бактерий — построить хромосомные карты бактерий (произвести картирование бактерий).
Трансформация и трансдукция
Мы познакомились с процессом удвоения ДНК, когда на одной цепи, как на матрице, выстраивается другая цепь. Однако в природе существуют процессы, связанные с изменением структуры ДНК, но эти изменения идут в других направлениях.
Трансформация – внесение в клетку генетической информации при помощи изолированной дезоксирибонуклеиновой кислоты (ДНК). Трансформация приводит к появлению у трансформированной клетки (трансформанта) и её потомства новых признаков, характерных для объекта — источника ДНК. Явление было открыто в 1928 английским учёным Ф. Гриффитом, наблюдавшим наследуемое восстановление синтеза капсульного полисахарида у пневмококков при заражении мышей смесью убитых нагреванием капсулированных бактерий и клеток, лишённых капсулы. Организм мыши в этих экспериментах играл роль своеобразного детектора, так как приобретение капсульного полисахарида сообщало клеткам, лишённым капсулы, способность вызывать смертельный для животного инфекционный процесс. В последующих экспериментах было установлено, что трансформация имеет место и в том случае, когда вместо убитых клеток к лишённым капсулы пневмококкам добавляли экстракт из разрушенных капсулированных бактерий. В 1944 О. Эйвери с сотрудниками (США) установил, что фактором, обеспечивающим трансформацию, являются молекулы ДНК. Эта работа — первое исследование, доказавшее роль ДНК как носителя наследственной информации.
Помимо пневмококков, трансформация обнаружена и изучена на некоторых других бактериях.
Трансформацию у бактерий рассматривают как сложный процесс, включающий следующие стадии:
– фиксация молекул ДНК клеткой-реципиентом;
– проникновение ДНК внутрь клетки;
– включение фрагментов трансформирующей ДНК в хромосому клетки-хозяина;
– формирование “чистых” трансформированных вариантов.
Фиксация ДНК происходит на особых участках клеточной поверхности (рецепторах), число которых ограничено. Связанная с рецепторами ДНК сохраняет чувствительность к действию добавленного в среду фермента дезоксирибонуклеазы, вызывающего её распад. Однако, спустя очень короткий срок (в пределах 1 мин) после фиксации, часть ДНК проникает в клетку. Бактериальные клетки одного и того же штамма резко различаются по проницаемости для ДНК. Клетки данной бактериальной популяции, способные включать чужеродную ДНК, называются компетентными. Число компетентных клеток в популяции незначительно и зависит от генетических особенностей бактерий и фазы роста бактериальной культуры. Развитие компетенции связывают с синтезом особого белка, обеспечивающего проникновение ДНК в клетку.
Средние размеры фрагментов ДНК, проникающих в клетку, составляют 5×106 дальтон. Поскольку в компетентную клетку может одновременно проникнуть ряд таких фрагментов, суммарная величина поглощённой ДНК может быть примерно равна размерам хромосомы клетки-хозяина. После проникновения в клетку двунитевой ДНК одна нить распадается до моно- и олигонуклеотидов, вторая — встраивается в хромосому клетки-хозяина путём её разрывов и воссоединений. Последующая репликация такой гибридной структуры приводит к выщеплению “чистых” клонов трансформантов, в потомстве которых закреплен признак, кодируемый включившейся ДНК.
Применение трансформации позволило провести генетический анализ бактерий, у которых не описано иных форм генетического обмена (конъюгации, трансдукции). Кроме того, трансформация – удобный метод для выяснения влияний на биологическую активность ДНК физических или химических изменений её структуры. Разработка метода у кишечной палочки позволила использовать для трансформации не только фрагменты бактериальной хромосомы, но и ДНК бактериальных плазмид и бактериофагов. Этот метод широко используется для внесения в клетку гибридной ДНК в исследованиях по генной инженерии.
Трансдукция (от лат. transductio — перемещение) – перенос генетического материала из одной клетки в другую с помощью вируса, что приводит к изменению наследственных свойств клеток-реципиентов. Явление трансдукции было открыто американскими учёными Д. Ледербергом и Н. Циндером в 1952. Особые бактериальные вирусы – умеренные фаги в процессе вегетативного размножения способны случайно захватывать и переносить в другие клетки любые участки ДНК лизируемых, то есть разрушаемых ими, бактерий (общая, или неспецифическая, трансдукция). Длина переносимого (трансдуцируемого) отрезка ДНК определяется размером белковой оболочки фаговой частицы и обычно не превышает 1-2% бактериального генома. Переносимый отрезок может содержать несколько генов. Поскольку вероятность такой сцепленной трансдукции зависит от расстояния между генами в молекуле ДНК, образующей хромосому бактерии, явление трансдукции широко используется при составлении генетических карт хромосом бактерий. Генетический материал фага в таких трансдуцирующих частицах отсутствует; поэтому, вводя ДНК в клетку, они не осуществляют все остальные функции фага: не размножаются, не лизогенизируют клетку и не наделяют её иммунитетом к фагу. Внесённый фрагмент может существовать в клетке в виде дополнительной генетического элемента, обладающего функциональной активностью. Поскольку такой фрагмент не способен воспроизводиться, при каждом клеточном делении он передаётся лишь в одну из дочерних клеток. За исключением этой клетки свойства всего остального потомства остаются без изменений (абортивная трансдукция). В дальнейшем фрагмент может быть либо разрушен, либо включен в хромосому бактерии, заменив в ней гомологичный участок ДНК. В последнем случае новые признаки, приобретённые клеткой-трансдуктантом, будут свойственны всему потомству этой клетки (полная трансдукция).
Существует группа бактериофагов, способных трансдуцировать лишь определённые гены, расположенные рядом с местом включения генома фага в хромосому бактерии при лизогенизации (ограниченная, или специфическая, трансдукция). Такие трансдуцирующие фаговые частицы, образующиеся в результате случайного нарушения точности процесса выхода профага из бактериальной хромосомы, содержат молекулу ДНК, состоящую из остатка фагового генома и фрагмента бактериального генома. В большинстве случаев они не могут самостоятельно размножаться или лизогенезировать бактерии из-за утраты части фагового генома (до 30%). Генетический материал трансдуцирующих частиц может сохраняться в клетке в автономном состоянии или в качестве профага включиться в ДНК бактерии. Однако в обоих случаях часть потомства восстанавливает исходные свойства из-за утраты профага. Стабильная трансдукция достигается только в случае включения бактериального фрагмента профага в геном бактерии в результате обмена на гомологичный участок хромосомы.
Эписомы (от эпи. и греч. sóma — тело) – генетические факторы, способные находиться в клетке либо в автономном (в цитоплазме) либо в интегрированном (включенными в хромосому) состоянии; представляют собой молекулы ДНК. К эписомам относятся геном умеренного фага лямбда, половой фактор F, некоторые R-факторы, сообщающие бактериям устойчивость к определённым лекарств, веществам, и некоторые др. Эписомы не являются обязательными компонентами клеток и могут переходить из одного состояния в другое, что зависит от вида клеток. Например, геном умеренного фага лямбда в клетках кишечной палочки может находиться в интегрированном либо в автономном состоянии, а в клетках возбудителя брюшного тифа – только в автономном состоянии. Находясь в автономном состоянии, большинство эписом ведут себя как типичные плазмиды. Ряд авторов видит в эписомах переходное звено между структурами, определяющими хромосомную и нехромосомную наследственность.
ДНК- диагностика
Организм человека является средой обитания для сотен видов бактерий и вирусов. С биологической точки зрения организм человека представляет собой целую систему сосуществующих организмов-симбионтов. Далеко не все из симбионтов патогенные. Без некоторых видов бактерий человек просто не способен существовать, их утрата или снижение количества является причиной развития ряда тяжелых заболеваний. Расшифровка геномов многих болезнетворных микроорганизмов с идентификацией всех белков поможет разработать методы предупреждения и лечения инфекционных болезней.
Для развития инфекционного процесса большое значение имеет генетический статус самого хозяина. Например, отдельные индивидуумы являются носителями вируса иммунодефицита, но СПИДом не болеют. У этих лиц имеется мутация в гене, кодирующем поверхностный белок, ответственный за попадание вируса внутрь лимфоидных клеток. Плотность белка на поверхности клеток снижена, вирус удерживается, но внутрь не попадает. Частота гомозигот по этой мутации среди жителей Европы составляет около 1%, они имеют выраженную устойчивость к ВИЧ-инфекциям. Более устойчивыми оказываются и гетерозиготные носители мутации, в российской популяции их частота достигает 13%.
Генетическая рекомбинация при трансформации. Трансдукция у бактерий. Общая и специфическая трансдукция. Использование трансформации и трансдукции для картирования генов. Трансдукция. Неспецифическая трансдукция. Специфическая трансдукция. Абортивная транс
Трансдукция — перенос бактериофагом в заражаемую клетку фрагментов генетического материала клетки, исходно содержавшей бактериофаг. Трансдуцирующий бактериофаг обычно переносит лишь небольшой фрагмент ДНК хозяина от одной клетки (донор) к другой (реципиент).
Выделено три типа трансдукции: неспецифическая (общая), специфическая и абортивная. В клетке, инфицированной бактериофагом, в ходе сборки дочерней популяции в головки некоторых фагов вместе с вирусной ДНК могут проникнуть фрагменты бактериальной ДНК или плазмиды. Вирусы ограничены в объёме генетического материала в соответствии с объёмом головки. Если ДНК бактериальной клетки расщепляется фагом в нетипичном месте, то чтобы освободить пространство для фрагмента хромосомной ДНК, некоторые участки вирусных ДНК «приносятся в жертву», что приводит к утере определённых их функций. При этом фаговая частица может стать дефектной. Количество аномальных фагов может достигать 0,3% всей дочерней популяции.
Образовавшийся фаг и есть частица, вызывающая неспецифическую (общую) трансдукцию. При такой форме трансдукции в клетки-реципиенты могут быть внесены практически любые гены.
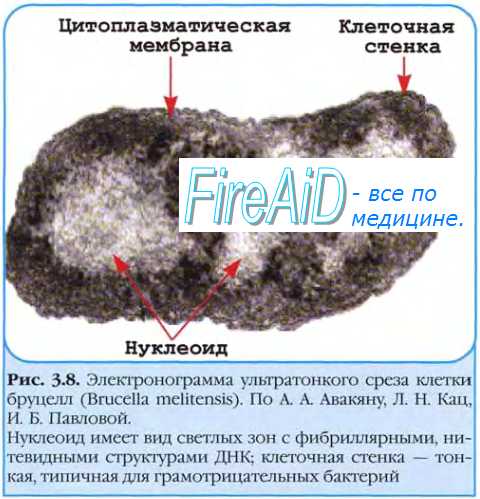
При неспецифической трансдукции фагом может быть перенесён любой фрагмент ДНК хозяина, а при специфической лишь строго определённые фрагменты ДНК. Наиболее известным примером специфической трансдукции служит трансдукция, осуществляемая фагом. Поскольку этот фаг при переходе в состояние профага включается в хромосому бактерий между генами, кодирующими синтез галактозы и биотина, именно эти гены он может переносить при транедукции. При абортивной трансдукции внесённый фрагмент ДНК донора не встраивается в генофор реципиента, а остаётся в цитоплазме, где его ДНК транскрибируется, но не реплицируется. Это приводит к тому, что при клеточном делении он передаётся только одной из дочерних клеток (то есть наследуется однолинейно) и затем теряется в потомстве.
Свойства трансдуцирующих фаговых частиц следующие:
• Частицы несут лишь часть ДНК фага, то есть не являются функциональными вирусами, а скорее ёмкостями, переносящими фрагменты бактериальной ДНК.
• Подобно прочим дефектным вирусам, частицы не способны к репликации.
• Трансдуцирующие фаги могут содержать какую-либо часть хромосомы хозяина с генами, дающими реципиентной бактерии некоторые преимущества (например, гены устойчивости к антибиотикам или гены, кодирующие способность к синтезу различных веществ). Подобное приобретение бактериями новых свойств получило название феномен лизогении.
• Феномен трансдукции может быть использован для картирования бактериальной хромосомы, если следовать тем же принципам, что и при картировании с использованием феномена трансформации.
Генетическая рекомбинация при трансформации. Трансдукция у бактерий. Общая и специфическая трансдукция. Использование трансформации и трансдукции для картирования генов
Трансформация бактерий – это перенос ДНК, изолированной из одних клеток в другие. Длинные фрагменты молекулы ДНК успешно поглощаются клеткой. Для того чтобы ДНК проникла в бактериальные клетки, они должны находиться в состоянии компетентности. Возникновению компетентности, приобретаемой лишь частью клеток культуры обычно в середине логарифмической стадии роста, способствует особый белок, который вырабатывается в ходе роста культуры. Сначала ДНК связывается с поверхностью компетентных клеток. ДНК, связанная с компетентными клетками, расщепляется специальными нуклеазами до фрагментов, которые проникают в клетку. После попадания в бактерию двуцепочечная ДНК превращается в одно цепочечную: одна нить ДНК деградирует. На заключительной стадии происходит интеграция одноцепочечного трансформирующего фрагмента с ДНК клетки-реципиента. При этом репликация не требуется, и включаемый фрагмент физически объединяется с ДНК реципиента. Весь процесс трансформации завершается в течение 10—30 мин. Частота трансформации разных бактерий составляет около 1 %. Для некоторых бактерий показана трансформация в естественных условиях, например в организме инфицированного животного — для Васcilus рпеитоniае, а также в условиях культуры — для Васcilus subtilis. Это означает, что трансформация — не экзотический прием генетического анализа, а естественный биологический процесс. В то же время в последние годы в связи с развитием генной инженерии широко применяется плазмидная, или векторная, трансформация, которая заключается во введении в клетки бактерий, а также эукариот генов, интегрированных в естественные или искусственные плазмиды. Трансдукцией называют перенос генов из одних бактериальных клеток в другие при помощи бактериофага. Это явление в 1951 г. открыл Н. Зиндер. Известны два основных типа бактериофагов — вирулентные и умеренные. Первые после их размножения в клетке бактерии приводят к ее лизису. Они существуют либо в вегетативном (размножение внутри клетки), либо в зрелом (метаболически инертное состояние вне клетки) состоянии. Умеренные бактриофаги обладают способностью быть в состоянии профага. Профагом был назван геном фага, который включается в бактериальную хромосому, после чего этот геном приобретает способность ауторепродуцироваться вместе с хромосомой бактерии. Такие бактерии, несущие профаг, называют лизогенными. Эти бактерии, хотя они и содержат в своей хромосоме профаг, не имеют в себе инфекционных фаговых частиц. Существование профага является временным. При индукции профаг покидает хромосому бактерий, переходит в вегетативное состояние и размножается. В этом случае в клетке возникают зрелые фаговые частицы, которые вызывают ее лизис. Этот процесс можно вызвать искусственно. Оказалось, что, когда профаг освобождается из хромосомы бактерии, переходя в стадию вегетативного существования, он может уносить с собой часть хромосомы бактерий. При следующем инфицировании гены из фрагмента хромосомы бактерии, унесенного фагом, оказываются способными внедряться в хромосому нового реципиента. Особый интерес имеет так называемая зиготическая индукция, при которой индуцирующим агентом является конъюгация с клеткой Hfr, при последней происходит проникновение хромосомы с профагом из клетки Hfr в нелизогенную клетку F. Обычно при трансдукции переносятся отдельные гены. Однако иногда переносимый сегмент хромосомы содержит два или несколько генов. Анализ таких случаев множественной трансдукции позволил создать карты участков хромосом бактерий. Существует несколько типов трансдукции: общая или неспецифическая – профаг способен включаться в разные места хромосомы бактерии. Это обеспечивает возможность переноса различных локусов из хромосомы хозяина в хромосому реципиента. Перенос генов при общей трансдукции может привести к двум различным состояниям трансдуктантов: а) полная трансдукция – привнесенный ген наследуется стабильно, т.к. интегрирует с хромосомой реципиента; б) абортивная трансдукция – внесенный фрагмент не реплицируется и передается по одной линии при размножении трансдуктантов; 2. специфическая – происходит рекомбинация между фаговой и хромосомной бактериальной ДНК, поэтому фаговые трансдуцирующие частицы обязательно содержат ДНК обоих типов. При использовании рекомбинации по многим генам оказалось возможным построить генетические карты хромосом для генов фагов Т4, Т2, фага лямбда и др. С помощью ряда приемов удалось показать соответствие генетических карт положению генов непосредственно в хромосомах вирусов. При интенсивном перемешивании раствора ДНК происходит разрыв молекул ДНК. Можно добиться режима, при котором молекула ДНК рвется примерно пополам. При заражении бактерий половинками молекул было показано, что они несут в себе половину генетической карты. М. Мезельсон и Дж. Уэйгл показали наличие совпадения расстояния между генами на карте и размером хромосом фага лямбда, используя передачу изотопов при рекомбинировании ДНК фага. Эти генетические карты оказались замкнутыми в круг. Генетические карты кольцевых хромосом изучены для ряда вирусных частиц.
29. Закономерности нехромосомного наследования, отличие от хромосомного наследования. Методы изучения: реципрокные, возвратные и поглощающие скрещивания, метод трансплантации, биохимические методы.
Наследование, определяемое хромосомами, получило название ядерного или хромосомного. В тех же случаях, когда материальной основой наследования являются элементы цитоплазмы, оно называется нехромосомным или цитоплазматическим. Поскольку и у растений, и у животных яйцеклетка содержит много цитоплазмы, а мужская гамета ее, как правило, почти лишена, следует ожидать, что цитоплазматическое наследование, в отличие от хромосомного, должно осуществляться по материнской линии. Цитоплазматическое наследование не может характеризоваться такими строгими количественными закономерностями, как ядерное. Например, воздействие повышенной температурой на яйца самок наездника (Habrobracon juglandis) до оплодотворения приводит к изменению окраски тела у их потомства. В ряде случаев наследование признаков связано с особенностями цитоплазмы, возникающими в процессе индивидуального развития организма либо под влиянием факторов внешней среды (онтогенетическая или фенотипическая предетерминация), либо под влиянием генотипа (генотипическая предетерминация). В этом случае наследование некоторых признаков по материнской линии. Онтогенетическая предетерминация. Обусловлена изменениями в цитоплазме, возникающими в ней под влиянием определенных внешних факторов. Обычно такие изменения нестойки, например, воздействие повышенной температурой на яйца самок наездника до оплодотворения приводит к изменению окраски тела у их потомства. Генотипическая предетерминация цитоплазмы происходит под влиянием генотипа материнского организма. Яркий пример – наследование направления завитка раковины у пресноводных гермафродитных моллюсков Limnea. Большинство из них – перекрестно оплодотворяющиеся формы, но некоторые из них способны к самооплодотворению. У этих моллюсков встречаются два типа закручивания раковины: против часовой стрелки (левозакрученные) и по ходу часовой стрелки (правозакрученные). При этом типе наследования фенотип потомков соответствует генотипу матери, а не генотипу зигот, из которых они развиваются. Данный признак предопределяется генотипом материнского организма в цитоплазме яйца в процессе его развития. В данном случае свойства цитоплазмы детерминированы действием хромосомных генов, а не элементами самой цитоплазмы, то есть здесь действует механизм хромосомного наследования, который изменяет цитоплазму яйцеклетки еще до оплодотворения. Реципрокное скрещивание, система из двух скрещиваний — прямого и обратного. При Р. с. каждый из генотипически различных родительских типов А и В используется дважды — один раз в качестве материнской и другой раз в качестве отцовской форм (♀А ´♂В и ♀В ´♂А). Различия между реципрокными гибридами могут быть вызваны влиянием материнского организма, цитоплазматической наследственности, сцепленными с полом генами. Метод возвратного скрещивания состоит в получении потомства в ряду поколений от скрещивания гетерозиготы (детей гомозиготных родителей, генетически отличающихся друг от друга) с одним из исходных гомозиготных родителей. Смысл подобного скрещивания – замена гена (генов какого-либо комплекса) одной инбредной линии на гаплотип другой. В результате получается конгенная линия, отличающаяся от исходной только по этому гену (генам этого комплекса). Поглотительное скрещивание, преобразовательное скрещивание, один из видов скрещивания, применяемый для коренного улучшения малопродуктивных пород высокопродуктивными. Простое П. с. заключается в спаривании животных двух пород (улучшаемой и улучшающей) для получения помесей, которых затем в ряде поколений спаривают с производителями улучшающей породы до получения животных желательного типа. Высокопродуктивных помесей 4—5—6-го поколений (высококровных), отвечающих типу улучшающей породы, разводят «в себе» (см. Разведение «в себе»), что иногда заканчивается созданием новой породы. П. с., в котором участвуют несколько улучшающих пород, называемых сложным. П. с. — наиболее быстрый и эффективный способ массового улучшения малопродуктивного скота, а также преобразования пород с.-х. животных (например, грубошёрстных пород овец в тонкорунных и полутонкорунных). Скорость преобразования и улучшения пород зависит от степени наследственных различий между животными скрещиваемых пород, степени наследственной устойчивости (консолидации) пород, тщательности отбора и подбора среди помесей, а также условий кормления и содержания помесного молодняка.
Методы трансплантации ядер В нашей стране Б.В. Конюховым и Е.С. Платоновым в 1985 г. был разработан метод менее травматического переноса ядер методом микроманипуляции. Он протекает в два этапа: сначала тонкой микропипеткой прокалывают зоны пеллюцида и плазматической мембраны и извлекают пронуклеусы, а затем другой пипеткой, большего диаметра (12 мкм) в то же отверстие вводят диплоидное ядро донора. В этом случае меньше травмируется цитоплазма зиготы и транспортируемое ядро донора. Трансплантация ядер может осуществляться и другим способом, с использованием цитохалазинов (веществ, синтезируемых грибами). Цитохалазин В разрушает структуру микрофиламентов и способствует уникальному расположению ядра. Ядро остается соединенным с клеткой тоненьким стебельком цитоплазмы. При центрифугировании этот мостик разрывается, образуются безъядерные клетки (цитопласты) и кариопласты, представляющие собой ядра, окруженные тонким слоем цитоплазмы и цитоплазматической мембраной. Цитопласты отделяют от интактных клеток в градиенте плотности. Они сохраняют способность прикрепляться к поверхности культурального сосуда и могут быть использованы для слияния с кариопластами других клеток с целью получения жизнеспособной клетки. Биохимические методы направлены на выявление биохимического фенотипа организма. Биохимические показатели (первичный белковый продукт гена, накопление патологических метаболитов внутри клетки) отражают сущность болезни более адекватно, чем клинические симптомы. С помощью биохимических методов описано более 1000 врожденных болезней обмена веществ. К закономерностям цитоплазматического наследования следует отнести: 1) передачу признаков по материнской линии; 2) отсутствие строгих количественных закономерностей расщепления. Принципами цитоплазматической наследственности являются: 1) дискретная детерминация признаков; 2) относительное постоянство плазмогенов; 3) множественность идентичных плазмогенов. Итак, понятие наследования и наследственности следует различать, по надо помнить, что в конечном итоге материальная и функциональная преемственность между поколениями обеспечивается всем и самовоспроизводящимися структурами клетки: ядерными и цитоплазматическими.
studopedia.org – Студопедия.Орг – 2014-2020 год. Студопедия не является автором материалов, которые размещены. Но предоставляет возможность бесплатного использования (0.001 с) . 

