Гмк гистология. Нормальная физиология. Потенциал действия гладких мышц сосудов
Гмк гистология. Нормальная физиология. Потенциал действия гладких мышц сосудов
Мембранные потенциалы в гладких мышцах. Величина мембранного потенциала гладкомышечного волокна зависит от состояния мышцы в данный момент. В норме в состоянии покоя внутриклеточный потенциал обычно составляет около -50-60 мВ, т.е. он примерно на 30 мВ менее отрицателен, чем в скелетной мышце.
Потенциалы действия в унитарных гладких мышцах. В гладких унитарных мышцах (например, во внутренних органах) потенциалы действия осуществляются тем же путем, как и в скелетных мышцах. В норме они не возникают во многих, если не в большинстве гладких мышц мультиу-нитарного типа, что обсуждается в следующем разделе. Существуют две формы потенциалов действия висцеральных гладких мышц: (1) спай-ковые потенциалы; (2) потенциалы действия с плато.
Спайковые потенциалы. В большинстве типов унитарных гладких мышц возникают типичные спайковые потенциалы, подобные тем, которые характерны для скелетных мышц. Длительность этих потенциалов действия составляет 10-50 мсек. Потенциалы действия этого типа могут возбуждаться многими путями, например при электрической стимуляции, действии гормонов на гладкую мышцу под влиянием медиаторов, выделяющихся из нервных волокон, при растяжении или в результате спонтанной генерации в самом мышечном волокне, что обсуждается далее.
Потенциалы действия с плато. На рисунке показан потенциал действия с плато гладкой мышцы. Начало этого потенциала действия подобно началу типичного спайкового потенциала. Однако реполяризация задерживается на период от нескольких сотен до 1000 мсек (1 сек). Наличие плато важно для поддержания длительного сокращения, характерного для некоторых типов гладких мышц, например мочеточника, матки в определенных условиях и некоторых кровеносных сосудов.

Значение кальциевых каналов в генерации потенциала действия гладких мышц. Мембрана гладкомышечных клеток содержит гораздо больше кальциевых электроуправляемых каналов, чем мембрана волокон скелетных мышц, и незначительное количество электроуправляемых натриевых каналов. В связи с этим участие натрия в генерации потенциала действия в большинстве гладких мышц незначительно. Вместо этого за развитие потенциала действия главным образом отвечает ток ионов кальция внутрь волокна. Это происходит по тому же принципу саморегенерации, как и при использовании натриевых каналов в нервных волокнах и волокнах скелетных мышц. Однако кальциевые каналы открываются во много раз медленнее, чем натриевые каналы, и остаются открытыми гораздо дольше.
Именно с этим главным образом связано длительное плато потенциалов действия некоторых гладкомышечных волокон.
Другой важной особенностью входа ионов кальция в клетки во время развития потенциала действия является их непосредственное влияние на сократительный механизм гладкой мышцы, в результате которого возникает сокращение. Следовательно, кальций одновременно решает две задачи.
Медленноволновые потенциалы в унитарных гладких мышцах и спонтанная генерация потенциалов действия.
Некоторые гладкие мышцы самовозбуждаются. Это означает, что потенциалы действия в гладкомышечных клетках возникают сами, без внешнего стимула. Часто это сочетается с основным медленноволновым ритмом колебаний мембранного потенциала. Сама медленная волна — не потенциал действия, т. е. она не связана с процессом саморегенерации, который способен распространяться по мембране мышечного волокна. Медленные волны являются отличительной особенностью гладкомышечных волокон, составляющих мышечную массу.
Причина медленноволнового ритма неизвестна. Одно предположение связывает существование медленных волн с усилением и ослаблением выкачивания положительных ионов (преимущественно ионов натрия) наружу через мембрану мышечного волокна, т.е. мембранный потенциал становится более отрицательным, когда натрий выкачивается быстро, и менее отрицательным, когда натриевый насос становится менее активным. Другое предположение связывает это явление с ритмическим увеличением и снижением проводимости ионных каналов.
Значение медленных волн состоит в том, что при достаточной амплитуде они могут инициировать потенциалы действия. Сами медленные волны не могут вызвать мышечное сокращение, но когда пик отрицательного медленно-волнового потенциала с внутренней стороны мембраны поднимается в положительном направлении от -60 до примерно -35 мВ (примерный порог для возбуждения в большинстве висцеральных гладких мышц), потенциал действия развивается и распространяется по мышечной массе. В этом случае действительно возникает сокращение. На рис. 8-4Б показан этот эффект: на каждом пике медленной волны возникают потенциалы действия — один или более.
Повторные последовательности потенциалов действия вызывают ритмическое сокращение гладкомышечной массы, поэтому медленные волны называют пейсмекерными волнами.
Гмк гистология. Нормальная физиология. Потенциал действия гладких мышц сосудов
Сосуды – это важный компонент сердечно-сосудистой системы. Они участвуют не только в доставке крови и кислорода к тканям и органам, но и осущевтляют регуляцию этих процессов.
1. Отличия в структуре стенки артерий и вен.
У артерий толстая мышечная медия, выраженный эластический слой.
Стенка вен менее плотная и более тонкая. Наиболее выраженный слой – адвентиция.

2. Типы мышечных волокон.
Многоядерные скелетные поперечно-полосатые мышечные волокна (по сути состоят не из отдельных клеток, а из синцитиев).
Кардиомиоциты тоже относятся к поперечно-полосатой мускулатуре, однако в них волокна связаны между собой контактами – нексусами, это обеспечивает распространение возбуждения по миокарду при его сокращении.
Гладкомышечные клетки имеют веретеновидную форму, они одноядерные.

3. Электронномикроскопическоая структура гладкой мышцы.

Типы гладкомышечных клеток
- мультунитарные
- унитарные

4. Фенотип гладкомышечной клетки.
5. Щелевые контакты в гладкой мышце осуществляют передачу возбуждения от клетки к клетке в унитарном типе гладких мышц.

6. Сравнительное изображение трех типов мышц.

7. Потенциал действия гладких мышц сосудов.

8. Тонический и фазический тип сокращений гладких мышц.

9. Пейсмейкерная активность гладких мышц вен, стенки желудочно-кишечного тракта и т.д.

10. Кривые сокращений скелетной, сердечной и гладкой мышц.

11. Механизм сокращений скелетных мышц.
Гладкие мышцы
Эти мышцы образуют мышечные слои стенок желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток. Гладкие мышцы разделяются на две основные группы: мультиунитарные и унитарные. Мультиунитарные мышцы функционируют независимо друг от друга, и каждое волокно может иннервироваться отдельным нервным окончанием. Такие волокна обнаружены в ресничной мышце глаза, мигательной перепонке и мышечных слоях некоторых крупных сосудов, к ним относятся мышцы, поднимающие волосы. У унитарных мышц волокна настолько тесно переплетены, что их мембраны могуг сливаться, образуя электрические контакты (нексусы). При раздражении одного волокна за счет этих контактов ПД быстро распространяются на соседние волокна. Поэтому, несмотря на то, что двигательные нервные окончания расположены на небольшом числе мышечных волокон, в реакцию вовлекается вся мышца. Такие мышцы имеются в большинстве органов: пищеварительном тракте, матке, в мочеточниках.
Особенностью гладких мышц является их способность осуществлять медленные и длительные тонические сокращения. Медленные, ритмические сокращения гладких мышц желудка, кишечника, мочеточников и других органов обеспечивают перемещение содержимого этих органов. Длительные тонические сокращения гладких мышц обеспечивают функционирование сфинктеров полых органов, которые препятствуют выходу их содержимого.
Гладкие мышцы стенок кровеносных сосудов, особенно артерий и артериол, также находятся в состоянии постоянного тонического сокращения. Изменение тонуса мышц стенок артериальных сосудов влияет на величину их просвета и, следовательно, на уровень кровяного давления и кровоснабжения органов. Важным свойством гладких мышц является их пластичность, т. е. способность сохранять приданную им при растяжении длину. Скелетная мышца в норме почти не обладает пластичностью. При удалении растягивающего груза скелетная мышца быстро укорачивается, а гладкая остается растянутой. Высокая пластичность гладких мышц имеет большое значение для нормального функционирования полых органов. Например, пластичность мышц мочевого пузыря по мере его наполнения предотвращает избыточное повышение давления.
Сильное и резкое растяжение гладких мышц вызывает их сокращение, что обусловлено нарастающей при растяжении деполяризацией клеток, которая обеспечивает автоматию гладкой мышцы. Такое сокращение играет важную роль в авторегуляции тонуса кровеносных сосудов, а также способствует непроизвольному опорожнению переполненного мочевого пузыря в тех случаях, когда нервная регуляция отсутствует в результате повреждения спинного мозга.
В гладких мышцах тетаническое сокращение возникает при низкой частоте стимуляции. В отличие от скелетных, гладкие мышцы способны развивать спонтанные тетанообразные сокращения в условиях денервации и даже после блокады интрамуральных ганглиев. Такие сокращения возникают вследствие активности клеток, обладающих автоматией (пейсмекерных клеток), которые отличаются по электрофизиологическим свойствам от других мышечных клеток. В них появляются пейсмекерные потенциалы, деполяризующие мембрану до критического уровня, что вызывает возникновение потенциала действия.
Особенностью гладких мышц является их высокая чувствительность к медиаторам, которые оказывают на спонтанную активность пейсмекеров модулирующие влияния. При нанесении ацетилхолина на препарат мышцы толстой кишки частота ПД возрастает. Вызываемые ими сокращения сливаются, образуется почти гладкий тетанус. Чем выше частота ПД, тем сильнее сокращение. Норадреналин, напротив, гиперполяризует мембрану, снижая частоту ПД и величину тетануса.
Возбуждение гладкомышечных клеток вызывает повышение концентрации кальция в саркоплазме, что активирует сократительные структуры. Так же как сердечная и скелетная мышцы, гладкая мышца расслабляется при снижении концентрации ионов кальция. Расслабление гладких мышц происходит медленнее, так как удаление ионов кальция замедлено.
Физиологические особенности гладких мышц
Пластичность гладкой мышцы. Важным свойством гладкой мышцы является ее большая пластичность, т. е. способность сохранять приданную растяжением длину без изменения напряжения. Различие между скелетной мышцей, обладающей малой пластичностью, и гладкой мышцей с хорошо выраженной пластичностью легко обнаруживается, если их сначала медленно растянуть, а затем снять растягивающий груз. Скелетная мышца тотчас укорачивается после снятия груза. В отличие от этого гладкая мышца после снятия груза остается растянутой до тех пор, пока под влиянием какого-либо раздражения не возникнет ее активного сокращения.
Свойство пластичности имеет очень большое значение для нормальной деятельности гладких мышц стенок полых органов, например мочевого пузыря: благодаря пластичности гладкой мускулатуры стенок пузыря давление внутри него относительно мало изменяется при разной степени наполнения.
Функциональный синцитий. Существуют различные типы гладких мышц. В стенках большинства полых органов находятся гладкие мышечные волокна длиной 50—400 мкм и диаметром 2—10 мкм. Эти волокна очень тесно примыкают Друг к другу и потому при .рассмотрении их в микроскопе создается впечатление, что они переходят друг в друга и морфологически составляют единое целое. На этом основании .утверждали, что гладкие мышцы, как и сердечная мышца, имеют синцитиальное строение. Однако электронно-микроскопические исследования показали, что не существует мембранной и цитоплазматической непрерывности между отдельными волокнами гладких мышц: они отделены друг от друга межклеточными щелями, ширина которых может достигать 60— 150 нм. Несмотря на наличие этих щелей, гладкие мышцы функционируют так, как если бы они имели истинное синцитиальное строение. Это выражается в том, что потенциалы действия и медленные волны деполяризации беспрепятственно распространяются с одного волокна на другое. Ввиду этого понятие «синцитиальное строение» является в настоящее время скорее физиологическим, чем морфологическим. Синцитий — функциональное образование, в котором возбуждение может свободно переходить с одной клетки в другую. Двигательные нервные окончания расположены только на небольшом числе волокон гладких мышц. Однако вследствие беспрепятственного распространения возбуждения с одного волокна на другое вовлечение в реакцию всей мышцы может происходить, если нервный импульс поступает к. небольшому числу мышечных волокон.
В некоторых гладких мышцах, например в ресничной мышце глаза или радиальной мышце радужной оболочки, каждое волокно имеет самостоятельную иннервацию, подобно волокнам скелетной мышцы.
Электрическая активность гладких мышц. Потенциал покоя гладкомышечных волокон, обладающих автоматией, обнаруживает постоянные небольшие колебания. Величина его при внутриклеточном отведении равна 30—70 мВ (в среднем 50 мВ). Потенциал покоя гладких мышечных волокон, не обладающих автоматией, стабилен и равен 60— 70 мВ. В обоих случаях его величина меньше значения потенциала покоя скелетных мышц. Это, по-видимому, связано с тем, что мембрана гладких мышечных волокон в покое характеризуется относительно высокой проницаемостью для ионов Na + .
Потенциалы действия в гладких мышцах также несколько ниже, чем в скелетных. Превышение потенциала действия над величиной потенциала покоя наблюдается не всегда и составляет не больше 10—20 мВ. В гладких мышцах внутренних органов зарегистрированы потенциалы действия двух основных типов: пикоподобные потенциалы
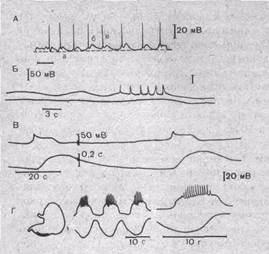
Рис.40. Электрическая и механическая активность различных гладких мышц.
А — изменение мембранного потенциала taenia coli морской свинки:а — потенциал покоя,б — вторичный ритм, в — пики; Б — потенциал действия и медленные колебания мембранного потенциала релаксационного типа в клетках собаки; В — потенциал действия типа плато в клетках гладкой мышцы аорты черепахи. Верхняя кривая — потенциал, нижняя — механическое напряжение сосудистой стенки; Г — спонтанная электрическая и механическая активность гладкомышеччых клеток желудка морской свинки (область отведения показана слева на схеме).
действия и потенциалы действия с выраженным плато. Длительность пикоподобных потенциалов действия варьирует от 5 до 80 мс. Пик, как правило, сопровождается следовой гиперполяризацией. Иногда наблюдается следовая деполяризация.
Потенциалы действия с выраженным плато зарегистрированы в гладких мышцах уретры, матки и некоторых’сосудов. Продолжительность плато 30—500 мс (рис. 40).
Ионный механизм возникновения потенциалов действия в гладких мышцах несколько отличается от такового в скелетных мышцах. Установлено, что деполяризация “мембраны, лежащая в основе потенциала действия в ряде гладких мышц, связана с активацией электровозбудимых кальциевых каналов. Следует подчеркнуть, что эти каналы проницаемы не только для ионов Са 2+ и некоторых двухвалентных катионов (Ва 2+ , Sr 2+ ), но и для ионов Na + . От «быстрых» натриевых каналов, обеспечивающих генерацию потенциалов действия в нервных и скелетно-мышечных волокнах, «медленные» кальциевые каналы отличаются не только своей ионной избирательностью, но также кинетикой процессов активации и инактивации и чувствительностью к блокаторам. Кальциевые каналы активируются и инактивируются значительно медленнее, чем натриевые; они не чувствительны к тетродотоксину, но эффективно блокируются изоптином (верапамилом), ионами Са 2+ , Мn 2+ и La 3+ . Изоптин применяют в медицинской практике для устранения или предупреждения спазма сосудов. .
Проведение возбуждения по гладкой мышце. В нервных и скелетных мышечных волокнах возбуждение распространяется посредством локальных электрических токов, возникающих между деполяризованным и соседними покоящимися участками клеточной мембраны. Этот же’механизм свойствен и волокнам гладких мышц. Однако в гладких мышцах потенциал действия, возникший в одном волокне (клетке), может распространяться на соседние волокна. Обусловлено это тем, что в мембранах клеток гладких мышц в области контактов с соседними клетками, так называемых нексусов, имеются участки относительно малого сопротивления, через которые петли тока, возникшие в одном волокне, легко проходят в соседние, вызывая деполяризацию их мембран. В этом отношении гладкая мышца отличается от скелетной и сходна с сердечной, которая также представляет собой функциональный синцитий. Между сердечным и гладкомышечным синцитием имеются некоторые важные различия. В сердце достаточно возбудить только одну клетку, чтобы это возбуждение распространилось на всю мышцу. В гладких же мышцах потенциал действия, возникший в одном участке, распространяется от него лишь на определенное расстояние, которое оказывается тем большим, чем сильнее приложенный стимул.
Другая существенная особенность гладких мышц заключается в том, что распространяющийся потенциал действия возникает в них только в том случае, если прило