Органы движения эукариотической клетки. Эукариотические клетки. Роль мутаций в эволюции
ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ
ПРОИСХОЖДЕНИЕ
Ископаемые останки клеток эукариотического типа обнаружены в породах, возраст которых не превышает 1,0—1,4 млрд. лет. Более позднее возникновение, а также сходство в общих чертах их основных биохимических процессов (самоудвоение ДНК, синтез белка на рибосомах) заставляют думать о том, что эукариотические клетки произошли от предка, имевшего прокариотическое строение.
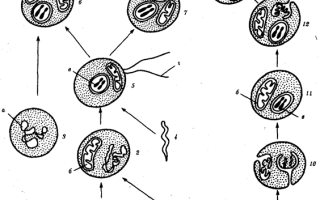
Наиболее популярна в настоящее время симбиотическая гипотеза происхождения эукариотических клеток, согласно которой (рис. 1.4) основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот, способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов — аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней.
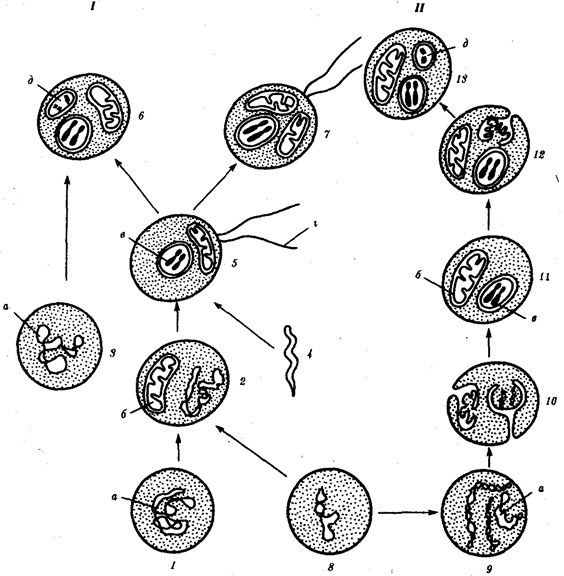
Рис. 1.4. Происхождение эукариотической клетки согласно симбиотической (I) и инвагинационной (II) гипотезам:
1 — анаэробный прокариот (клетка-хозяин), 2 — прокариоты, имеющие митохондрии, 3 — сине-зеленая водоросль (презумптивный хлоропласт), 4 —сиирохетообразная бактерия (презумпгивный жгутик), 5 — примитивный эукариот со жгутиком, 6 — растительная клетка, 7 — животная клетка со жгутиком, 8 — аэробный прокариот (презумптивная митохондрия), 9 — аэробный прокариот (клетка-родоначальница согласно гипотезе II), 10 —инвагинации клеточной оболочки, давшие ядро и митохондрии, 11 — примитивный эукариот 12 — впячивание клеточной оболочки, давшее хлоропласт, 13 — растительная клетка; а—ДНК прокариотической клетки, б — митохондрия, в —ядро эукариотической клетки, г —жгутик, д — хлоропласт
Сходное происхождение предполагают для жгутиков, предками которых служили симбионты-бактерии, имевшие жгутик и напоминавшие современных спирохет. Приобретение клеткой жгутиков имело наряду с освоением активного способа движения важное следствие общего порядка. Предполагают, что базальные тельца, которыми снабжены жгутики, могли эволюционировать в центриоли в процессе возникновения механизма митоза.
Способность зеленых растений к фотосинтезу обусловлена присутствием в их клетках хлоропластов. Сторонники симбиотической гипотезы считают, что симбионтами клетки-хозяина, давшими начало хлоропластам, послужили прокариотические синезеленые водоросли.
Серьезным доводом в пользу симбиотического происхождения митохондрий, центриолей и хлоропластов является то, что перечисленные органеллы имеют собственную ДНК. Вместе с тем белки бациллин и тубулин, из которых состоят жгутики и реснички соответственно современных прокариот и эукариот, имеют различное строение. У бактерий не найдено также структур со свойственной жгутикам, ресничкам, базальным тельцам и центриолям эукариотических клеток комбинацией микротрубочек: «9 + 2» или «9 + 0».
Внутриклеточные мембраны гладкой и шероховатой цитоплазматической сети, пластинчатого комплекса, пузырьков и вакуолей рассматривают как производные наружной мембраны ядерной оболочки, которая способна образовывать впячивания.
Центральным и трудным для ответа является вопрос о происхождении ядра. Предполагают, что оно также могло образоваться из симбионта-прокариота. Увеличение количества ядерной ДНК, во много раз превышающее в современной эукариотической клетке ее количество в митохондрий или хлоропласте, происходило, по-видимому, постепенно путем перемещения групп генов из геномов симбионтов. Нельзя исключить, однако, что ядерный геном формировался путем наращивания генома клетки-хозяина (без участия симбионтов).
Согласно инвагинационной гипотезе, предковой формой эукариотической клетки был аэробный прокариот (рис. 1.4). Внутри такой клетки-хозяина находилось одновременно несколько геномов, первоначально прикреплявшихся к клеточной оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран.
Инвагинационная гипотеза хорошо объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Однако она не может ответить на вопрос, почему биосинтез белка в хлоропластах и митохондриях в деталях соответствует таковому в современных прокариотических клетках, но отличается от биосинтеза белка в цитоплазме эукариотической клетки.
История показала, что эволюционные возможности клеток эукариотического типа несравнимо выше, чем прокариотического. Ведущая роль здесь принадлежит ядерному геному эукариот, который во много раз превосходит по размерам геном прокариот. Количество генов у бактерии и в клетке человека, например, соотносится как 1: (100-1000). Важные отличия заключаются в диплоидности эукариотических клеток благодаря наличию в ядрах двух комплектов генов, а также в многократном повторении некоторых генов. Это расширяет масштабы мутационной изменчивости без угрозы резкого снижения жизнеспособности, эволюционно значимым следствием чего является образование резерва наследственной изменчивости.
При переходе к эукариотическому типу усложняется механизм регуляции жизнедеятельности клетки, что на уровне генетического материала проявилось в увеличении относительного количества регуляторных генов, замене кольцевых «голых» молекул ДНК прокариот хромосомами, в которых ДНК соединена с белками. В итоге стало возможным считывать биологическую информацию по частям с разных групп генов в разном их сочетании в различных типах клеток и в разное время. В бактериальной клетке, напротив, одновременно считывается до 80—100% информации генома. В клетках взрослого человека в разных его органах транскрибируется от 8—10% (печень, почка) до 44% (головной мозг) информации. Использованию биологической информации частями принадлежит исключительная роль в эволюции многоклеточных организмов, так как именно это позволяет разным группам клеток специализироваться по различным функциональным направлениям.
Большое значение при переходе к многоклеточности имело наличие у эукариотических клеток эластичной оболочки, что необходимо для образования устойчивых клеточных комплексов.
Среди цитофизиологических особенностей эукариот, увеличивающих их эволюционные возможности, необходимо назвать аэробное дыхание, которое также послужило предпосылкой для развития многоклеточных форм. Интересно, что сами эукариотические клетки появились на Земле после того, как концентрация O2 в атмосфере достигла 1% (точка Пастера). Названная концентрация является необходимым условием аэробного дыхания.
В условиях усложнения генетического аппарата эукариот, увеличения суммарного количества ДНК и распределения ее по хромосомам трудно переоценить значение возникновения в эволюции митоза как механизма воспроизведения в поколениях генетически сходных клеток.
Появление вследствие эволюционных преобразований митоза такого способа деления клеток, как мейоз, дающего возможность сохранить постоянство хромосом в ряду поколений, наилучшим образом решило проблему размножения многоклеточных организмов. Связанный с мейозом переход к половому размножению усилил эволюционную роль комбинативной изменчивости, способствовал увеличению скорости эволюции.
Благодаря отмеченным особенностям за 1 млрд. лет эволюции эукариотический тип клеточной организации дал широкое разнообразие живых форм от одноклеточных простейших до млекопитающих и человека.
Происхождение эукариот
Расцвет эукариот на Земле начался около 1 млрд лет назад, хотя первые из них появились намного раньше (возможно 2,5 млрд лет назад). Происхождение эукариот могло быть связано с вынужденной эволюцией прокариотических организмов в атмосфере, которая стала содержать кислород.
Симбиогенез — основная гипотеза происхождения эукариот
Существует несколько гипотез о путях возникновения эукариотических клеток. Наиболее популярная — симбиотическая гипотеза (симбиогенез). Согласно ей, эукариоты произошли в результате объединения в одной клетке разных прокариот, которые сначала вступили в симбиоз, а затем, все более специализируясь, стали органоидами единого организма-клетки. Как минимум симбиотическое происхождение имеют митохондрии и хлоропласты (пластиды вообще). Произошли они от бактериальных симбионтов.
Клеткой-хозяином мог быть относительно крупный анаэробный гетеротрофный прокариот, похожий на амебу. В отличие от других, он мог приобрести способность питаться путем фаго- и пиноцитоза, что позволяло ему захватывать других прокариот. Они не все переваривались, а снабжали хозяина продуктами своей жизнедеятельности). В свою очередь, получали от него питательные вещества.
Митохондрии произошли от аэробных бактерий и позволили клетке-хозяину перейти к аэробному дыханию, которое не только намного эффективней, но и облегчает существование в атмосфере, содержащей достаточно большое количество кислорода. В такой среде аэробные организмы получают преимущество над анаэробными.
Позже в некоторых клетках поселились похожие на ныне живущих синезеленых водорослей (цианобактерий) древние прокариоты. Они стали хлоропластами, дав начало эволюционной ветви растений.
Кроме митохондрий и пластид симбиотическое происхождение могут иметь жгутики эукариот. В них превратились симбионты-бактерии наподобие современных спирохет, имеющих жгутик. Считается, что в последствии из базальных тел жгутиков произошли центриоли, столь важные структуры для механизма клеточного деления эукариот.
Эндоплазматическая сеть, комплекс Гольджи, пузырьки и вакуоли могли произойти от наружной мембраны ядерной оболочки. С другой точки зрения, некоторые из перечисленных органелл могли возникнуть путем упрощения митохондрий или пластид.
Во многом неясным остается вопрос происхождения ядра. Могло ли оно также образоваться из прокариота-симбионта? Количество ДНК в ядре современных эукариот во много раз превышает его количество в митохондриях и хлоропластах. Возможно часть генетической информации последних со временем переместилась в ядро. Также в процессе эволюции происходило дальнейшее увеличение размера ядерного генома.
Кроме того в симбиотической гипотезе происхождения эукариот не все так однозначно с клеткой-хозяином. Им мог и не быть один вид прокариот. Используя методы сравнения геномов, ученые делают вывод, что клетка-хозяин близок к археям, при этом сочетает в себе признаки архей и ряда неродственных групп бактерий. Отсюда можно сделать вывод, что появление эукариот происходило в сложном сообществе прокариот. При этом процесс скорее всего начался с метаногенной археи, вступавшей в симбиоз с другими прокариотами, что было вызвано необходимостью обитания в кислородной среде. Появление фагоцитоза способствовало притоку чужих генов, а ядро образовалось для защиты генетического материала.
Молекулярный анализ показал, что различные белки эукариот происходят от разных групп прокариот.
Доказательства симбиогенеза
В пользу симбиотического происхождения эукариот говорит то, что митохондрии и хлоропласты имеют собственную ДНК, причем кольцевую и не связанную с белками (также обстоит дело у прокариот). Однако в генах митохондрий и пластид есть интроны, чего нет у прокариот.
Пластиды и митохондрии не воспроизводятся клеткой с нуля. Они образуются из ранее существующих таких же органелл путем их деления и последующего роста.
В настоящее время существуют амебы, у которых нет митохондрий, а вместо них есть бактерии симбионты. Также есть простейшие, сожительствующие с одноклеточными водорослями, выполняющими в клетке-хозяине роль хлоропластов.
Инвагинационная гипотеза происхождения эукариот
Кроме симбиогенеза существуют и другие взгляды на происхождение эукариот. Например, инвагинационная гипотеза. Согласно ей, предком эукариотической клетки был не анаэробный, а аэробный прокариот. К такой клетке могли прикрепляться другие прокариоты. Потом их геномы объединялись.
Ядро, митохондрии и пластиды возникли путем впячивания и отшнуровывания участков клеточной мембраны. В эти структуры попадала чужеродная ДНК.
Усложнение генома происходило в процессе дальнейшей эволюции.
Инвагинационная гипотеза происхождения эукариот хорошо объясняет наличие двойной мембраны у органелл. Однако она не объясняет, почему система биосинтеза белка в хлоропластах и митохондриях сходна с прокариотической, в то время как таковая в ядерно-цитоплазматическом комплексе имеет ключевые отличия.
Причины эволюции эукариот
Все разнообразие жизни на Земле (от простейших до покрытосеменных и млекопитающих) дали клетки эукариотического, а не прокариотического типа. Возникает вопрос, почему? Очевидно, ряд особенностей, возникших у эукариот, существенно повысили их эволюционные возможности.
Во-первых, у эукариот есть ядерный геном, который во много раз превосходит количество ДНК у прокариот. При этом эукариотические клетки диплоидны, кроме этого в каждом гаплоидном наборе определенные гены многократно повторяются. Все это обеспечивает, с одной стороны, большие масштабы для мутационной изменчивости, а с другой — уменьшает угрозу резкого снижения жизнеспособности в результате вредной мутации. Таким образом, эукариоты, в отличие от прокариот, обладают резервом наследственной изменчивости.
Эукариотические клетки имеют более сложный механизм регуляции жизнедеятельности, у них существенно больше различных регуляторных генов. Кроме того, молекулы ДНК образовали комплексы с белками, что позволило наследственному материалу упаковываться и распаковываться. Все вместе это дало возможность считывать информацию частями, в разных сочетаниях и количестве, в разное время. (Если в клетках прокариот транскрибируется почти вся информация генома, то в эукариотических клетках обычно менее половины.) Благодаря этому эукариоты могли специализироваться, лучше приспосабливаться.
У эукариот появились митоз, а затем и мейоз. Митоз позволяет воспроизводить генетически сходные клетки, а мейоз сильно увеличивает комбинативную изменчивость, что ускоряет эволюцию.
Большую роль в процветании эукариот сыграло приобретенное их предком аэробное дыхание (хотя оно есть и у многих прокариот).
На заре своей эволюции эукариоты обзавелись эластичной оболочкой, обеспечивавшей возможность фагоцитоза, и жгутиками, позволившими им двигаться. Это дало возможность эффективней питаться.
Строение эукариотической клетки
Строение клетки
Строение клеток
Все клеточные формы жизни на земле можно разделить на два надцарства на основании строения составляющих их клеток — прокариоты (доядерные) и эукариоты (ядерные). Прокариотические клетки — более простые по строению, по-видимому, они возникли в процессе эволюции раньше. Эукариотические клетки — более сложные, возникли позже. Клетки, составляющие тело человека, являются эукариотическими.
Несмотря на многообразие форм организация клеток всех живых организмов подчинена единым структурным принципам.
Живое содержимое клетки — протопласт — отделено от окружающей среды плазматической мембраной, или плазмалеммой. Внутри клетка заполнена цитоплазмой, в которой расположены различные органоиды и клеточные включения, а также генетический материал в виде молекулы ДНК. Каждый из органоидов клетки выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом.
Прокариотическая клетка


Строение типичной клетки прокариот: капсула, клеточная стенка, плазмалемма, цитоплазма, рибосомы, плазмида, пили, жгутик, нуклеоид.
Прокариоты (от лат. pro — перед, до и греч. κάρῠον — ядро, орех) — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды.
Эукариотическая клетка Эукариоты (эвкариоты) (от греч. ευ — хорошо, полностью и κάρῠον — ядро, орех) — организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты — митохондрии, а у водорослей и растений — также и пластиды.
Строение хромосом

Схема строения хромосомы в поздней профазе — метафазе митоза. 1—хроматида; 2—центромера; 3—короткое плечо; 4—длинное плечо.
Хромосо́мы (др.-греч. χρῶμα — цвет и σῶμα — тело) — нуклеопротеидные структуры в ядре эукариотической клетки (клетки, содержащей ядро), которые становятся легко заметными в определённых фазах клеточного цикла (во время митоза или мейоза). Хромосомы представляют собой высокую степень конденсации хроматина, постоянно присутствующего в клеточном ядре. Исходно термин был предложен для обозначения структур, выявляемых в эукариотических клетках, но в последние десятилетия всё чаще говорят о бактериальных хромосомах. В хромосомах сосредоточена большая часть наследственной информации.
Морфология хромосом лучше всего видна в клетке на стадии метафазы. Хромосома состоит из двух палочкообразных телец – хроматид. Обе хроматиды каждой хромосомы идентичны друг другу по генному составу.
Хромосомы дифференцированы по длине. Хромосомы имеют центромеру или первичную перетяжку, две теломеры и два плеча. На некоторых хромосомах выделяют вторичные перетяжки и спутники. Движение хромосомы определяет Центромера, которая имеет сложное строение.
ДНК центромеры отличается характерной последовательностью нуклеотидов и специфическими белками. В зависимости от расположения центромеры различают акроцентрические, субметацентрические и метацентрические хромосомы.
Как говорилось выше, некоторые хромосомы имеют вторичные перетяжки. Они, в отличие от первичной перетяжки (центромеры), не служат местом прикрепления нитей веретена и не играют никакой роли в движении хромосом. Некоторые вторичные перетяжки связаны с образованием ядрышек, в этом случае их называют ядрышковыми организаторами. В ядрышковых организаторах расположены гены, ответственные за синтез РНК. Функция других вторичных перетяжек еще не ясна.
У некоторых акроцентрических хромосом есть спутники — участки, соединенные с остальной частью хромосомы тонкой нитью хроматина. Форма и размеры спутника постоянны для данной хромосомы. У человека спутники имеются у пяти пар хромосом.
Концевые участки хромосом, богатые структурным гетерохроматином, называются теломерами. Теломеры препятствуют слипанию концов хромосом после редупликации и тем самым способствуют сохранению их целостности. Следовательно, теломеры ответственны за существование хромосом как индивидуальных образований.
Хромосомы, имеющие одинаковый порядок генов, называют гомологичными. Они имеют одинаковое строение (длина, расположение центромеры и т. д.). Негомологичные хромосомы имеют разный генный набор и разное строение.
Исследование тонкой структуры хромосом показало, что они состоят из ДНК, белка и небольшого количества РНК. Молекула ДНК несет отрицательные заряды, распределенные по всей длине, а присоединенные к ней белки — гистоны заряжены положительно. Этот комплекс ДНК с белком называют хроматином. Хроматин может иметь разную степень конденсации. Конденсированный хроматин называют гетерохроматином, деконденсированный хроматин — эухроматином. Степень деконденсации хроматина отражает его функциональное состояние. Гетерохроматиновые участки функционально менее активны, чем эухроматиновые, в которых локализована большая часть генов. Различают структурный гетерохроматин, количество, которого различается в разных хромосомах, но располагается он постоянно в околоцентромерных районах. Кроме структурного гетерохроматина существует факультативный гетерохроматин, который появляется в хромосоме при сверхспирализации эухроматических районов. Подтверждением существования этого явления в хромосомах человека служит факт генетической инактивации одной Х-хромосомы в соматических клетках женщины. Его суть заключается в том, что существует эволюционно сформировавшийся механизм инактивации второй дозы генов, локализованных в Х-хромосоме, вследствие чего, несмотря на разное число Х-хромосом в мужском и женском организмах, число функционирующих в них генов уравнено. Максимально конденсирован хроматин во время митотического деления клеток, тогда его можно обнаружить в виде плотных хромосом
Размеры молекул ДНК хромосом огромны. Каждая хромосома представлена одной молекулой ДНК. Они могут достигать сотен микрометров и даже сантиметров. Из хромосом человека самая большая — первая; ее ДНК имеет общую длину до 7 см. Суммарная длина молекул ДНК всех хромосом одной клетки человека составляет 170 см.
Несмотря на гигантские размеры молекул ДНК, она достаточно плотно упакована в хромосомах. Такую специфическую укладку хромосомной ДНК обеспечивают белки гистоны. Гистоны располагаются по длине молекулы ДНК в виде блоков. В один блок входит 8 молекул гистонов, образуя нуклеосому (образование, состоящее из нити ДНК, намотанной вокруг октамера гистонов). Размер нуклеосомы около 10 нм. Нуклеосомы имеют вид нанизанных на нитку бусинок. Нуклеосомы и соединяющие их участки ДНК плотно упакованы в виде спирали, на каждый виток такой спирали приходится шесть нуклеосом. Так формируется структура хромосомы.
Наследственная информация организма строго упорядочена по отдельным хромосомам. Каждый организм характеризуется определенным набором хромосом (число, размеры и структура), который называется кариотипом. Кариотип человека представлен двадцатью четырьмя разными хромосомами (22 пары аутосом, Х- и Y-хромосомы). Кариотип — это паспорт вида. Анализ кариотипа позволяет выявлять нарушения, которые могут приводить к аномалиям развития, наследственным болезням или гибели плодов и эмбрионов на ранних стадиях развития.
Длительное время полагали, что кариотип человека состоит из 48 хромосом. Однако в начале 1956 г. было опубликовано сообщение, согласно которому число хромосом в кариотипе человека равно 46.
Хромосомы человека различаются по размеру, расположению центромеры и вторичных перетяжек. Впервые подразделение кариотипа на группы было проведено в 1960 г. на конференции в г. Денвере (США). В описание кариотипа человека первоначально были заложены два следующих принципа: расположение хромосом по их длине; группировка хромосом по расположению центромеры (метацентрические, субметацентрические, акроцентрические).
Точное постоянство числа хромосом, их индивидуальность и сложность строения свидетельствуют о важности выполняемой ими функции. Хромосомы выполняют функцию основного генетического аппарата клетки. В них в линейном порядке расположены гены, каждый из которых занимает строго определенное место (локус) в хромосоме. В каждой хромосоме много генов, но для нормального развития организма необходим набор генов полного хромосомного набора.
Строение и функции ДНК
ДНК — полимер, мономерами которой являются дезоксирибонуклеотиды. Модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. Дж. Уотсоном и Ф. Криком (для построения этой модели они использовали работы М. Уилкинса, Р. Франклин, Э. Чаргаффа).
Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т.е. представляет собой двойную спираль (исключение — некоторые ДНК-содержащие вирусы имеют одноцепочечную ДНК). Диаметр двойной спирали ДНК — 2 нм, расстояние между соседними нуклеотидами — 0,34 нм, на один оборот спирали приходится 10 пар нуклеотидов. Длина молекулы может достигать нескольких сантиметров. Молекулярный вес — десятки и сотни миллионов. Суммарная длина ДНК ядра клетки человека — около 2 м. В эукариотических клетках ДНК образует комплексы с белками и имеет специфическую пространственную конформацию.
Мономер ДНК — нуклеотид (дезоксирибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов. Пиримидиновые основания ДНК (имеют в составе своей молекулы одно кольцо) — тимин, цитозин. Пуриновые основания (имеют два кольца) — аденин и гуанин.
Моносахарид нуклеотида ДНК представлен дезоксирибозой.
Название нуклеотида является производным от названия соответствующего основания. Нуклеотиды и азотистые основания обозначаются заглавными буквами.
2.4. Строение про– и эукариотической клеток.
Любая клетка представляет собой систему: все ее компоненты взаимосвязаны, взаимозависимы и взаимодействуют друг с другом; нарушение деятельности одного из элементов данной системы ведет к изменениям и нарушениям работы всей системы.
Совокупность клеток образует ткани, различные ткани образуют органы, а органы, взаимодействуя и выполняя общую функцию, образуют системы органов.
Любая система обладает определенной структурой, уровнем сложности и основана на взаимодействии элементов, которые ее составляют.
Особенности строения эукариотических и прокариотических клеток:


Строение эукариотических клеток.




Функции эукариотических клеток.
Клетки одноклеточных организмов осуществляют все функции, характерные для живых организмов – обмен веществ, рост, развитие, размножение; способны к адаптации.
Клетки многоклеточных организмов дифференцированы по строению, в зависимости от выполняемых ими функций. Эпителиальные, мышечные, нервные, соединительные ткани формируются из специализированных клеток.
Тематические задания
А1. К прокариотическим организмам относится
А2. Клеточная мембрана выполняет функцию
1) синтеза белка
2) передачи наследственной информации
4) фагоцитоза и пиноцитоза
А3. Укажите пункт, в котором строение названной клетки совпадает с ее функцией
1) нейрон – сокращение
2) лейкоцит – проведение импульса
3) эритроцит – транспорт газов
4) остеоцит – фагоцитоз
А4. Клеточная энергия вырабатывается в
4) аппарате Гольджи
А5. Исключите из предложенного списка лишнее понятие
А6. Исключите из предложенного списка лишнее понятие
4) крахмальные зерна
А7. Хромосомы клетки выполняют функцию
1) биосинтеза белка
2) хранения наследственной информации
3) формирования лизосом
4) регуляции обмена веществ
В1. Выберите из предложенного списка функции хлоропластов
1) образование лизосом
2) синтез глюкозы
5) выделение кислорода
6) клеточное дыхание
В2. Выберите особенности строения митохондрий