Оксигемоглобин: понятие, механизм образования, кривая диссоциации и её сдвиги. Факторы, влияющие на кривую диссоциации оксигемоглобина Диссоциация оксигемоглобина увеличивается при условии
Факторы, влияющие на кривую диссоциации оксигемоглобина
Форма кривой диссоциации НbО2 обусловлена главным образом реакционноспособностью гемоглобина, однако сродство крови к кислороду может измениться под действием других факторов, как правило, приводящим к увеличению или уменьшению наклона кривой диссоциации без изменения ее S–образной формы. Такое влияние оказывают температура, рН, напряжение СO2 и некоторые другие факторы, роль которых возрастает при патологических состояниях.
Влияние температуры. Равновесие реакции оксигенации гемоглобина (как и большинства химических реакций вообще) зависит от температуры. При понижении температуры наклон кривой диссоциации оксигемоглобина увеличивается, а при ее повышении – снижается
Таблица. Параметры дыхательных газов и рН для артериальной и венозной крови у здоровых молодых людей в покое
Влияние рН и PСO2. Форма кривой диссоциации оксигемоглобина в значительной степени зависит от содержания в крови ионов Н + .При снижении рН, т. е. подкислении крови, сродство гемоглобина к кислороду уменьшается и угол наклона кривой диссоциации оксигемоглобина уменьшается. рН крови тесно связан с напряжением в ней СO2 (рСO2): чем выше РO2 , тем ниже рН. Очевидно, сам диоксид углерода оказывает на диссоциацию оксигемоглобина специфическое действие.
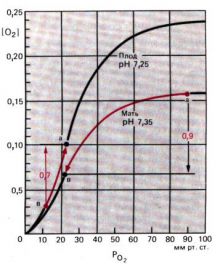
Поглощение O2 идет одновременно с выделением СO2, поэтому по мере насыщения гемоглобина кислородом кривая диссоциации оксигемоглобина сдвигается влево. По мере того как венозная кровь (точка в; pO2 = 40 мм рт. ст., РCO2 = 46 мм рт. ст.), насыщаясь кислородом, превращается в артериальную (точка а;PO2 = 95 мм рт. ст., РCO2 = 40мм рт. ст.), сродство гемоглобина к кислороду постоянно увеличивается. В результате, хотя перенос кислорода осуществляется путем диффузии, скорость этой диффузии несколько возрастает.
Поскольку одновременно с выходом кислорода из крови в нее поступает СO2, кривая диссоциации оксигемоглобина смещается вправо. Снижение сродства гемоглобина к кислороду приводит к еще большему падению содержания оксигемоглобина, и в результате кислород поступает в ткани при относительно высоком рО2 в капилляре.
Сродство гемоглобина к кислороду зависит также от содержания в эритроцитах катионов. Необходимо отметить и влияние патологических сдвигов рН: при повышении рН (алкалозе) захват кислорода в легких облегчается, но его отдача в тканях затрудняется, а при снижении рН (ацидозе) наблюдается обратная картина. Наконец, значительный сдвиг кривой диссоциации оксигемоглобина влево имеет место при отравлении СО.
37. Способы определения величины плеврального давления учебник
Измерение давления в плевральной полости: (1) прямой метод – прокол грудной стенки и введение в плевральную полость иглы, связанной с манометром; (2) непрямой метод – измерение давления с помощью зонда, введенного в пищевод (внутрипищеводное давление соответствует давлению в плевральной полости). В конце выдоха давление = -3-6 мм рт.ст (объем легкого уменьшился, ЭТЛ уменьшилась), а в конце вдоха давление = -6-9 мм рт.ст (объем легкого увеличился, ЭТЛ увеличилась).
38. Спирометрия, спирография. Работы 3.17, 18– стр.102, 104
(1) дыхательный объем (ДО) – объем воздуха, который мы вдыхаем (и выдыхаем) во время одного спокойного вдоха (и выдоха) – 500 мл. Определяется методом спирометрии.
(2) резервный объем вдоха (РОвд) – объем воздуха, который мы можем вдохнуть после спокойного вдоха – 2000 мл. Определяется методом спирометрии.
(3) резервный объем выдоха (РОвыд) – объем воздуха, который мы можем выдохнуть после спокойного выдоха – 1500 мл. Определяется методом спирометрии.
(4) остаточный объем (ОО) – объем воздуха, который остается в легких после максимального выдоха – 1000 мл. Определяется методом разведения индикатора (гелий).
ЛЕГОЧНЫЕ ЕМКОСТИ (каждая емкость состоит из 2-х и более объемов)
(1) жизненная емкость легких (ЖЕЛ) – максимальный объем воздуха, который мы можем выдохнуть после максимально глубокого вдоха (ДО + РОвд + РОвыд) = 4-5 литров (значение: показатель общего физического развития). Определяется методом спирометрии.
(2) емкость вдоха – максимальный объем воздуха, который мы можем вдохнуть после спокойного выдоха (ДО + РОвд). Определяется методом спирометрии.
(3) функциональная остаточная емкость (ФОЕ) – объем воздуха, который остается в легких после спокойного выдоха (РОвыд + ОО) = 2500 мл (значение: показатель состояния эластической тяги легких. При снижении ЭТЛ этот показатель увеличивается). Определяется методом плетизмографии, разведения индикатора.
(4) общая емкость легких (ОЕЛ) – объем воздуха, который находится в легких после максимально глубокого вдоха (сумма всех 4-х объемов) = 5-6 литров. Определяется методом плетизмографии, разведения индикатора.
39.Показатели вентиляции: минутный объем дыхания, альвеолярная вентиляция, максимальная вентиляция легких. . Работа 3.21– стр.110 + учебник
Минутный объем дыхания (МОД) – объем воздуха, который проходит через легкие за минуту. МОД = ДО (дыхательный объем) х ЧД (частота дыхания) = 6-8 л/мин
Альвеолярная вентиляция (АВ) – объем воздуха, который проходит через альвеолярное пространство за минуту и участвует в газообмене.
АВ = (ДО – ОМП) х ЧД, где ОМП – объем мертвого пространства (150 мл – объем дыхательных путей, в котором не происходит газообмен). Например, АВ = (500 – 150) х 12 = 4200 мл/мин
Максимальная вентиляция легких (МВЛ) – максимальный объем воздуха, который может пройти через легкие за минуту (при максимально возможной глубине и частоте дыхания). Показывает резервные возможности дыхательного аппарата. Достигает 180 л/мин. (Исследование проводится 10-15 сек).

40. Оксигемометрия, пульсоксиметрия . Работа 3.25– стр.114
Пульсоксиметрия — методика определения количества кислорода, связанного с гемоглобином, в артериальной крови. К каждой молекуле гемоглобина может присоединится до четырех молекул кислорода. Средний процент насыщения молекул гемоглобина является кислородной сатурацией крови. 100% сатурация означает, что к каждая молекула гемоглобина в исследуемом объеме крови переносит четыре молекулы кислорода.
Принцип работы пульсоксометра основан на дифференцированном поглощении света с разной длинной волны гемоглобином в зависимости от степени насыщения кислородом.
Пульсоксиметор состоит из источника света двух длин волн (660 нм «красный» и 940 нм «инфракрасный»), фотоприемника, процессора, монитора.

- Периферический датчик изучает «красный» и «инфракрасный» свет.
- Кровь поглощает излучение (степень зависит от сатурации кислорода).
- Оставшийся световой поток улавливается фотоприемником.
- Полученные данные обрабатываются в процессорном блоке и выводятся на экран монитора.
1. 
41. Пневмотахометрия и пик-флоуметрия, индекс Тиффно . Работы 3.19, 20– стр.107, 108
Пневмотахометрия– определение объемной скорости движения воздуха (л/сек) через датчик во время (а) форсированного вдоха и (б) форсированного выдоха. При увеличении сопротивления дыхательных путей пневмотахометрические показатели уменьшаются.
Пикфлоуметрия(англ. Peak Flow) – метод функциональной диагностики для определения пиковой объемной скорости форсированного выдоха. Иными словами данный метод помогает оценить, с какой скоростью человек может выдохнуть воздух, и таким образом оценить степень обструкции (сужения) дыхательных путей.
Для данного метода исследования существует специальный прибор – пикфлоуметр, который представляет собой компактную трубочку с градуированной шкалой.
Процедура проводится в положении сидя (или стоя). Сначала следует сделать несколько спокойных вдохов и выдохов, после чего делается глубокий вдох, мундштук пикфлоуметра плотно обхватывается губами и производится глубокий форсированный выдох. При этом следует держать аппарат строго параллельно поверхности пола. За каждый сеанс требуется сделать не менее 3 выдохов через некоторые промежутки времени (2-3 мин.), и выбрать максимальное значение.

При достижении им максимального результата (которые должны быть приближены к норме) его умножают на коэффициент 0.8 – например, если максимальный показатель пикфлоуметрии 400 л/мин, то 400 умножают на 0,8. Получаем 320 л/мин. Значение измерения выше этого показателя будет относиться к «зеленой зоне» – то есть нормальному уровню проходимости дыхательных путей. «Желтая зона» – это максимальный показатель исследования, умноженный на коэффициент 0,5. То есть 400 умножаем на 0,5 и получаем 200 мл/мин (это будет нижней границей желтой зоны). В этом случае границами «желтой зоны» будут значения от 200 л/мин до 320 л/мин.


Определение объема форсированного выдоха (проба Тиффно) за первую секунду (ОФВ1) – во время спирометрического исследования пациент должен сделать максимальный вдох, задержать дыхание на вдохе, а потом как можно быстрее выдохнуть. За первую секунду форсированного выдоха в норме он должен выдохнуть 70% от форсированной жизненной емкости легких. При увеличении сопротивления дыхательных путей этот показатель уменьшается.
Сенсорные системы

42. Определение остроты зрения. Работа 6.1– стр.232
43. Аккомодационный рефлекс. Значение. Работа 6.2– стр.233
44. Зрачковый рефлекс Физиологическое значение. Работа 6.9– стр.239
45. Исследование цветового зрения. Работа 6.5– стр.236
46. Определение световой и темновой адаптации зрения (адаптометрия) учебник
Кривая диссоциации оксигемоглобина, факторы, влияющие на ее ход.
Способность гемоглобина реагировать с кислородом характеризует кривая диссоциации оксигемоглобина (КДО). При ее построении на оси абсцисс видкладують РО2 (мм.рт.ст), по оси ординат – содержание оксигемоглобина в крови. Строят КДО минимум по двум точкам:
при РО2 = 100 мм.рт.ст. гемоглобин насыщен кислородом на 98%;
при РО2 = 60 мм.рт.ст. насыщение О2 составляет 90%;
при РО2 = 26 мм.рт.ст. насыщения гемоглобина кислородом – 50%.
при РО2 = 0, насыщения гемоглобина = 0%.
Характерно, что при высоком РО2 гемоглобин легко взаимодействует с кислородом – образование HbO2 (верхняя, полога – “горизонтальная” часть кривой). Снижение РО2 со 100 до 60 мм.рт.ст мало влияет на образование НbO2 – его концентрации уменьшается лишь на 8%. Это означает, что снижение давления кислорода в альвеолах до 60 мм.рт.ст мало повлияет на транспорт кислорода кровью, хотя напряжение кислорода в плазме будет снижаться пропорционально снижению давления О2 в альвеолах. Благодаря такой особенности хода КДО, мы можем, например, подниматься в горы – несмотря на существенное снижение атмосферного давления, снабжение тканей кислородом сохраняется на должном уровне.
Когда парциальное давление О2 в атмосфере высокий, реакция
сдвинута в сторону образования оксигемоглобина. В условиях целостного организма такие умовистворюються при прохождении крови капиллярами легких.
Снижение Ро2 ниже 60 мм.рт.ст. сопровождается значительным снижением HbO2 в крови – он активно распадается с образованием гемоглобина и свободного кислорода. В условиях целостного организма это происходит в тканях (уровень Ро2 составляет 50-20 мм.рт.ст.). И чем активнее функционирует ткань, тем ниже в ней уровень О2 – усиленная диссоциация HbO2 с высвобождением молекулярного кислорода, который утилизируется тканями. То есть, в этих условиях реакция взаимодействия кислорода и гемоглобина сдвинута в сторону диссоциации оксигемоглобина.
Итак, S-образная форма кривой диссоциации оксигемоглобина отражает компромисс между необходимостью активно связывать кислород в атмосфере с высоким парциальным давлением его (капилляры легких) и легко отдавать кислород в атмосфере с низким парциальным давлением (капилляры тканей).
Родство кислорода и гемоглобина и ход КДО зависят от нескольких факторов. Некоторые из них снижают эту родство; при этом гемоглобин легче отдает кислород, а КДО сдвигается вправо. Видно, что при смещении КДО справа при том же уровне парциального давления кислорода распадается больше оксигемоглобина. К факторам, сдвигают КДО справа относятся:
– Накопление углекислоты (взаимодействует с глобиновой частью Нb – снижение его сродства к кислороду)
– Накопление ионов водорода (протонов – снижение рН) – протоны также взаимодействуют с глобина – снижение сродства кислорода и Hb;
Все эти цинникы влияют на оксигемоглобин в тканях, активпо работают – именно там накапливаются углекислота, протоны, повышается температура (через усиленный метаболизм). Это приводит к уменьшению сродства Hb и О2 – усиление диссоциации оксигемоглобина – усиленное образование молекулярного кислорода, который необходим тканям, активно функционируют.
Факторы, влияющие на кривую диссоциации оксигемоглобина
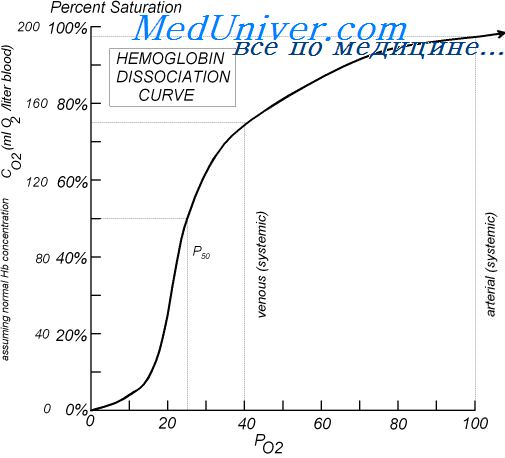
На процесс присоединения кислорода к гемоглобину влияют следующие факторы: концентрация ионов водорода, напряжение углекислого газа, температура, концентрация 2,3-дифосфоглице-рата (2,3-ДФГ). Их суммарный эффект на взаимодействие гемоглобина с кислородом отражает величина P50 — значение напряжения кислорода, при котором гемоглобин насыщен на 50% (рис. 22-23). Изменение каждого из факторов способно смещать кривую диссоциации вправо (увеличение P50) или влево (уменьшение Р.™). Сдвиг вправо вызывает снижение сродства гемоглобина к кислороду, вытесняет кислород из связи с гемоглобином и увеличивает количество кислорода, доступного тканям; сдвиг влево дает обратный эффект. В норме P50 составляет 26,6 мм рт. ст. (3,4 кПа).
Увеличение концентрации ионов водорода в крови снижает связывание гемоглобина с кислородом (эффект Бора).Форма кривой диссоциации оксигемоглобинатакова, что этот эффект более выражен в венозной крови, чем в артериальной (рис. 22-23); данный феномен облегчает осво-
бождение кислорода в тканях, практически не сказываясь на потреблении кислорода (в отсутствие тяжелой гипоксии).
Влияние напряжения CO2 на сродство гемоглобина к кислороду имеет важное физиологическое значение; вместе с тем оно вторично по отношению к увеличению концентрации ионов водорода, которая возрастает с увеличением PCO2. Высокое содержание CO2 в венозном сегменте капилляров, снижая сродство гемоглобина к кислороду, облегчает освобождение кислорода в тканях; наоборот, низкое содержание CO2 в легочных капиллярах вновь увеличивает сродство гемоглобина к кислороду, облегчая поглощение кислорода из альвеол.
2,3-ДФГ, побочный продукт гликолиза (шунт Rapoport-Luebering), накапливается при анаэробном метаболизме. Хотя эффект 2,3-ДФГ на гемоглобин теоретически благоприятен для организма (уменьшается сродство гемоглобина к кислороду и облегчается освобождение O2 в тканях.— Примеч. ред.), его физиологическое значение невелико. 2,3-ДФГ, тем не менее, играет важную компенсаторную роль при хронической анемии и существенно влияет на транспортную функцию гемоглобина донорской крови при гемотрансфузиях (гл. 29).

Рис. 22-22. Кривая диссоциации оксигемоглобина у здорового взрослого человека. (С разрешения. Из: West J. В. Respiratory Physiology: The Essentials, 3rd ed. Williams & Wilkins, 1985.)
Аномальные лиганды и аномальные формы гемоглобина
Окись углерода (СО), цианиды, азотная кислота и аммиак moitt связываться с гемоглобином в местах соединения с кислородом. Они вытесняют кислород и смещают кривую диссоциации влево. Угарный газ отличается особенной активностью: его сродство к гемоглобину в 200-300раз выше, чем у кислорода. СО связывается с гемоглобином, образуя карбоксигемоглобин, что снижает кислородную емкость гемоглобина и нарушает освобождение кислорода в тканях.
При окислении железа тема до трехвалентной формы образуется метгемоглобин. В редких случаях нитраты, нитриты, сульфаниламиды и другие лекарственные средства могут вызывать сильную метгемоглобинемию. Метгемоглобин неспособен связывать кислород, до тех пор пока он не будет восстановлен с помощью фермента метгемо-глобинредуктазы; кроме того, метгемоглобин смещает кривую диссоциации оксигемоглобина влево. Метгемоглобинемия, как и отравление угарным газом, снижает кислородную емкость крови и нарушает высвобождение кислорода в тканях. Метилено-вый синий и аскорбиновая кислота способствуют восстановлению метгемоглобина в гемоглобин.
Аномальные формы гемоглобина возникают в результате изменений в составе белковых субъединиц. Каждый вариант имеет собственные характеристики связывания с кислородом. Наиболее распространенные аномальные формы гемоглобина включают фе-тальный гемоглобин, гемоглобин A2, гемоглобин при серповидно-клеточной анемии (гл. 29).
Содержание кислорода в крови
Общее содержание кислорода в крови равно сумме физически растворенного и связанного с гемоглобином кислорода. Связывание кислорода с гемоглобином никогда не достигает теоретического максимума, поэтому считают, что 1 г гемоглобина может связать приблизительно 1,31 мл кислорода. Содержание кислорода в крови (С, от англ, content — содержание) выражается следующим уравнением:
Содержание кислорода (в 100 мл крови) = = [(0,003 мл О2/ЮО мл крови/мм рт. ст.) х PO2] + + (SO2X HbX 1,31 мл/1 OO мл крови),
где Hb — концентрация гемоглобина (г/100 мл крови), a SO2 — насыщение гемоглобина кислородом (S, от англ, saturation — насыщенеие) при данном PO2.

Рис. 22-23.Сдвиги кривой диссоциации оксигемоглобина при изменениях рН, температуры тела и концентрации 2,3-дифосфоглицерата (2,3-ДФГ) в эритроцитах
Используя эту формулу и величину Hb, равную 15 г/100 мл, можно рассчитать содержание O2 в артериальной pi смешанной венозной крови, а также артериовенозную разницу по кислороду (при SaO2 – 97,5 % и SvO2 = 75 %):
= 19,5 мл О2/100 мл крови;
CvO2-(0,003 х 40)+ (0,75 х 15 х 1,31) =
Транспорт кислорода
Транспорт кислорода зависит как от дыхания, так и от кровообращения (гл. 19). Общая доставка кислорода (DO2; от англ, delivery — доставка) к тканям равна произведению содержания кислорода в артериальной крови и сердечного выброса:
Заметим, что содержание кислорода в артериальной крови зависит как от РлО2, так и от концентрации гемоглобина. Следовательно, недостаточная доставка кислорода может быть результатом низкого РлО2, низкой концентрации гемоглобина или низкого сердечного выброса. В норме расчет доставки кислорода выглядит так:
DO2 = 20 мл О2/1OO мл крови х 5000 мл /мин =
Уравнение Фикавыражает связь между потреблением O2, артериовенозной разницей по кислороду и сердечным выбросом:
Потребление O2 = VO2 = Qt x (CaO2 – CvO2). После преобразования получаем:
Следовательно, артериовенозную разницу можно рассматривать как меру адекватности доставки кислорода.
При нормальном потреблении кислорода около 250 мл/мин и сердечном выбросе 5000 мл/мин нормальная артериовенозная разница, согласно этому уравнению, составит 5 мл O2/!OO мл крови. Заметим, что при этом нормальный коэффициент экстракции O2 [(CaO2 – CvO2)/CaO2] составит 25 %, т. е. 5 мл/20 мл. Таким образом, в норме организм потребляет только 25 % кислорода, переносимого гемоглобином. Когда потребность в O2 превосходит возможность его доставки, то коэффициент экстракции становится выше 25 %. На-
оборот, если доставка O2 превышает потребность, то коэффициент экстракции падает ниже 25 %.
Если доставка кислорода снижена умеренно, потребление кислорода не изменяется благодаря увеличению экстракции O2 (насыщение гемоглобина кислородом в смешанной венозной крови снижается); в этом случае VO2 не зависит от доставки. По мере дальнейшего снижения DO2 достигается критическая точка, в которой VO2 становится прямо пропорционально DO2. Состояние, при котором потребление кислорода зависит от доставки, характеризуется прогрессирующим лак-тат-ацидозом (гл. 30), обусловленным клеточной гипоксией.
Кислородный резерв
Понятие кислородного резерва имеет большое значение в анестезиологии. Когда в результате апноэ прекращается поступление в организм кислорода, то в ходе клеточного метаболизма потребляется имеющийся кислородный резерв; после того как резерв исчерпан, развивается гипоксия и наступает смерть клеток. Теоретически нормальный кислородный резерв у взрослого человека составляет около 1500 мл. Он включает остатки кислорода в легких; кислород, находящийся в связи с гемоглобином и миоглобином; кислород, растворенный в жидкостях организма. К сожалению, высокое сродство гемоглобина к кислороду (сродство мио-глобина к кислороду еще выше), а также незначительное количество кислорода, физически растворенного в тканях, представляют собой очень малый резерв. Следовательно, основным источником кислорода является дыхательная смесь, находящаяся в легких в объеме, соответствующем ФОЕ (исходный объем при апноэ). Необходимо отметить, что только около 80 % этого объема может быть использовано.
При наступлении апноэ у больного, дышавшего перед этим атмосферным воздухом, в легких имеется примерно 480 мл кислорода (если FiO2 = 0,21 и ФОЕ = 2300 мл, то объем кислорода составит FiO2 X ФОЕ; 0,21 X 2300 мл = 480 мл). Метаболически активные ткани быстро используют этот резерв (предположительно со скоростью потребления кислорода); в течение 90 с развивается тяжелая гипоксемия. Возникновение гипоксемии можно отсрочить путем увеличения FiO2 перед апноэ. После дыхания чистым кислородом легкие содержат 2300 мл кислорода, что задерживат развитие гипоксемии после апноэ на 4-5 мин. Данная концепция лежит в основе проведения преоксигена-ции перед индукцией анестезии (гл. 5).
УГЛЕКИСЛЫЙ ГАЗ
CO2 транспортируется кровью в физически растворенном виде, в составе бикарбоната и в комплексе с белками в виде карбаминовых соединений (табл. 22-6). Сумма всех трех форм составляет общее содержание CO2 в крови, которое стандартным образом измеряется при анализе электролитов.
Физически растворенный CO2
CO2 растворяется в крови лучше, чем кислород, коэффициент его растворимости 0,031 ммоль/л/ мм рт. ст. при 37 0 C (0,067 мл/100 мл/мм рт. ст.)
Бикарбонат
В водных растворах CO2 медленно вступает в связь с водой, образуя бикарбонат:
В плазме в эту реакцию вступает менее 1 % растворенного CO2, тогда как в эритроцитах и в эндо-телиальных клетках имеется фермент карбо-ангидраза, который ее ускоряет. В результате бикарбонат представляет самую большую фракцию CO2 в крови (табл. 22-6). Ацетазоламид, будучи ингибитором карбоангидразы, может нарушать доставку CO2 от тканей к альвеолам.
В венозных сегментах капилляров большого круга кровообращения CO2 поступает в эритроциты, где трансфомируется в бикарбонат, который диффундирует из эритроцитов в плазму. Для поддержания электрического равновесия из плазмы в эритроциты перемещаются ионы Cl”. В легочных капиллярах происходит обратный процесс: ионы Cl” выходят из эритроцитов, а бикарбонат поступает в них для превращения в CO2, который диффундирует в альвеолы. Перемещение ионов
Cl” носит название хлоридного сдвига,или сдвига Гамбургера.
Карбаминовые соединения
CO2 может реагировать с аминогруппами белков согласно реакции:
При физиологических значениях рН только небольшое количество CO2 переносится в этой форме, главным образом, в комплексе с гемоглобином (карбаминогемоглобин). Сродство дезоксигениро-ванного гемоглобина (дезоксигемоглобина) к CO2 в 3,5 раза выше, чем у оксигемоглобина. Увеличение сродства крови к CO2 при ее деоксигенации часто называют эффектом Холдейна(табл. 22-6). В норме PCO2 существенно не влияет на фракцию CO2, которая транспортируется в виде карбами-ногемоглобина.
Влияние гемоглобинового буфера на транспорт CO2
Эффект Холдейна отчасти обусловлен буферными свойствами гемоглобина (гл. 30). При нормальном рН гемоглобин может выполнять роль буфера за счет высокого содержания гистидина. Кроме того, кислотно-основные свойства гемоглобина зависят от степени его оксигенации:
После высвобождения кислорода в тканевых капиллярах молекула гемоглобина начинает вести себя подобно основанию; связывая ионы водорода, гемоглобин смещает равновесие СО2-бикарбонат преимущественно в сторону образования бикарбоната:
ТАБЛИЦА 22-6. Транспорт СО2(из расчета на 1 л цельной крови)
Оксигемоглобин: понятие, механизм образования, кривая диссоциации и её сдвиги. Факторы, влияющие на кривую диссоциации оксигемоглобина Диссоциация оксигемоглобина увеличивается при условии
Кривые диссоциации оксигемоглобина справедливы для нормальной крови со средними показателями. Однако существует ряд факторов, которые могут сдвигать эту кривую в одну или другую сторону. На рисунке видно, что при некотором закислении крови со снижением рН от нормального уровня 7,4 до 7,2 кривая диссоциации смещается в среднем на 15% вправо, а повышение уровня рН от нормального уровня 7,4 до 7,6 смещает кривую на такое же расстояние влево.
Кроме изменений рН известны и другие факторы, которые могут сдвигать кривую диссоциации. Назовем три, действие которых сдвигает кривую вправо: (1) повышение концентрации двуокиси углерода; (2) повышение температуры крови; (3) повышение концентрации 2,3-дифосфоглицерата — метаболически важного фосфата, который в зависимости от метаболических условий присутствует в крови в разных концентрациях.
Повышение снабжения тканей кислородом в случаях, когда двуокись углерода и ионы водорода сдвигают кривую диссоциации оксигемоглобина. Эффект Бора. Сдвиг кривой диссоциации оксигемоглобина в ответ на повышение содержания двуокиси углерода и ионов водорода в крови имеет существенное влияние, выражающееся в ускорении высвобождения кислорода из крови в тканях и увеличении оксигенации крови в легких. Это называют эффектом Бора и объясняют его следующим образом.
При прохождении крови через ткань двуокись углерода диффундирует из клеток ткани в кровь. В результате в крови увеличивается Ро2, а затем концентрации угольной кислоты (Н2СО3) и ионов водорода. Эти изменения сдвигают кривую диссоциации оксигемоглобина вправо и вниз, уменьшая сродство кислорода к гемоглобину, и в результате увеличивается выход кислорода в ткани.
При диффузии двуокиси углерода из крови в альвеолы происходят процессы обратного направления — в результате в крови снижаются Рсо2 и концентрация ионов водорода, сдвигая кривую диссоциации оксигемоглобина влево и вверх. При этом’ значительно увеличивается количество кислорода, связывающегося с гемоглобином при любом существующем уровне альвеолярного Ро2, что увеличивает транспорт кислорода к тканям.
Сдвиг кривой диссоциации оксигемоглобина под влиянием дифосфоглицерата. Нормальное содержание ДФГ в крови вызывает постоянный небольшой сдвиг кривой диссоциации оксигемоглобина вправо. В случае гипоксического состояния, продолжающегося более нескольких часов, концентрация ДФГ в крови значительно возрастает, и кривая диссоциации оксигемоглобина сдвигается вправо еще больше.
В присутствии такой концентрации ДФГ кислород в тканях высвобождается при Ро2, превышающем нормальный уровень на 10 мм рт. ст., поэтому в некоторых случаях такой механизм с участием ДФГ может оказаться важным для адаптации к гипоксии, особенно если причиной гипоксии является уменьшение в ткани кровотока.
Сдвиг кривой диссоциации во время физической нагрузки. Во время физической нагрузки некоторые факторы вызывают значительный сдвиг кривой диссоциации оксигемоглобина вправо, поэтому активные, выполняющие физическую работу мышечные волокна получают дополнительное количество кислорода. В свою очередь, работающие мышцы высвобождают большое количество двуокиси углерода; это в совокупности с действием некоторых других кислот, высвобождающихся мышцами, повышает концентрацию ионов водорода в крови капилляров мышц.
Кроме того, во время работы температура мышцы часто повышается на 2-3°С, что может еще больше увеличивать доставку кислорода мышечным волокнам. Все эти факторы вызывают значительный сдвиг кривой диссоциации оксигемоглобина в крови капилляров мышц вправо. Сдвиг вправо означает высвобождение кислорода гемоглобином в мышце при достаточно высоком уровне Ро2 (40 мм рт. ст.) даже в случаях, когда из него уже высвободилось 70% кислорода. Сдвиг кривой в другую сторону показывает, что в легких присоединилось дополнительное количество кислорода из альвеолярного воздуха.